Frontiers in Neuroscience
Introduzione
Con il progresso della società e i cambiamenti nel ritmo di lavoro, un numero crescente di persone soffre di privazione del sonno. La privazione del sonno non solo danneggia la salute fisica e mentale dell’individuo, ma influisce anche seriamente sulle prestazioni lavorative, causando errori di lavoro e persino incidenti. Pertanto, comprendere il meccanismo della privazione del sonno che influenza la funzione cognitiva è di grande importanza per prevenire efficacemente gli effetti della privazione del sonno.
Studi precedenti hanno rivelato che la privazione del sonno può causare una serie di cambiamenti nell’umore di un individuo, nelle capacità cognitive, nelle prestazioni lavorative e nella funzione immunitaria (Choo et al., 2005). La mancanza di sonno interrompe la circolazione del corpo e influenza le capacità cognitive ed emotive degli individui (Raymond, 1988). Diversi studi hanno rivelato che la privazione del sonno compromette l’inibizione della risposta (Harrison e Horne, 1998; Muzur et al., 2002; Jennings et al., 2003). Ad esempio, dopo 36 ore di privazione del sonno, la capacità dell’individuo di sopprimere gli stimoli negativi è diminuita (Chuah et al., 2006). Studi di neuroimaging hanno suggerito che la privazione del sonno riduce il basso livello di capacità di elaborazione visiva di un individuo (Anderson e Platten, 2011; Ning et al., 2014). Inoltre, la privazione del sonno compromette l’ippocampo e potrebbe influenzare la memoria distruggendo la plasticità sinaptica (Cote et al., 2014). Thomas (2003) ha indicato che la mancanza di sonno ha ridotto il flusso sanguigno cerebrale e il tasso metabolico nel talamo, nella corteccia prefrontale e nella corteccia parietale (Géraldine et al., 2005). Jarraya e colleghi hanno scoperto che la privazione parziale del sonno ha influenzato significativamente le funzioni neuropsicologiche come la memoria istantanea verbale, l’attenzione e la vigilanza (Thomas, 2003). Inoltre, alcuni studi hanno rivelato che gli effetti cumulativi della privazione parziale del sonno potrebbero compromettere gravemente la funzione cognitiva e il comportamento (Van Dongen, 2004; Scott et al., 2006; Jarraya et al., 2013).
La memoria di lavoro è un sistema utilizzato per memorizzare ed elaborare informazioni e che è una funzione cognitiva con capacità limitata (Bartel et al., 2004). Inoltre, le informazioni memorizzate nel sistema di memoria di lavoro possono essere modificate dalla memoria a breve termine alla memoria a lungo termine attraverso la rivisitazione e altri metodi di memoria. La memoria di lavoro è la transizione tra sistemi di memoria a breve e a lungo termine, che è molto fondamentale nell’elaborazione dei messaggi umani (Miyake e Shah, 1999). Fornisce uno spazio di archiviazione temporaneo e le risorse necessarie per elaborare le informazioni, come la comprensione vocale, il ragionamento e l’apprendimento. È stato dimostrato che la privazione del sonno influisce prima sulla memoria di lavoro.
Studi precedenti hanno utilizzato il paradigma della memoria di lavoro n-back in partecipanti sottoposti a privazione del sonno e hanno scoperto che la mancanza di sonno induce una diminuzione dell’attività metabolica nella rete regionale del cervello, che viene principalmente effettuata elaborazione delle informazioni e inibizione della reazione (Baddeley, 2000; Zhang et al., 2019). La memoria di lavoro compromessa dopo la privazione del sonno è correlata all’attivazione della rete predefinita nelle attività (Chee e Chuah, 2008), che può essere correlata all’importante ruolo del talamo nella vigilanza corticale. Ad esempio, la privazione del sonno ha aumentato la connessione tra l’ippocampo, il talamo e la rete predefinita, che è stata spesso accompagnata da una maggiore sonnolenza soggettiva e da prestazioni peggiori della memoria di lavoro (Lei et al., 2015; Li et al., 2016). Studi sulla privazione del sonno identi?l’aumento della latenza e l’ampiezza ridotta della componente P3 sono stati associati a una prolungata sobrietà (Morris et al., 1992; Jones e Harrison, 2001; Panjwani et al., 2010). La diminuzione dell’onda P3 potrebbe riflettere una diminuzione dell’attenzione dei partecipanti e una riduzione del discernimento degli stimoli target (Koslowsky e Babkoff, 1992).
Tuttavia, pochi studi hanno fornito prove elettrofisiologiche per la compromissione della memoria di lavoro dopo la privazione del sonno. L’attività n-back è considerata un metodo comune per valutare la memoria di lavoro (Owen et al., 2005; Jaeggi et al., 2010). Zhang et al. progettato un compito di memoria di lavoro pronuncia due-back per esplorare l’alternativa messaggio diminuito di memoria di lavoro durante la privazione del sonno, ma pochi studi hanno utilizzato diversi tipi di attività di memoria di lavoro in un unico esperimento. Nel presente studio, abbiamo progettato diversi tipi di attività di memoria di lavoro (memoria di lavoro di pronuncia, memoria di lavoro spaziale e memoria di lavoro degli oggetti) per esplorare la compromissione della funzione cognitiva da parte del TSD e dei dati EEG dei partecipanti registrati in 2 punti temporali (baseline e 36 h-TSD). Tutti i compiti hanno adottato un paradigma 2-back. Questo studio ha valutato i cambiamenti nell’onda N2-P3 relativi alla memoria di lavoro durante il TSD e ha analizzato le caratteristiche temporali degli effetti della privazione del sonno sulla memoria di lavoro. I nostri risultati forniscono prove sperimentali per gli effetti della privazione del sonno sulla funzione cognitiva.
Materiali e metodi
Partecipanti
Sedici studenti maschi giovani, sani e destrorsi hanno partecipato a questo studio. Abbiamo reclutato partecipanti pubblicizzando nel campus. I partecipanti avevano tutti buone abitudini di sonno (PSQI<5). Tutti i partecipanti avevano un’età compresa tra 21 e 28 anni con un’età media di 23 anni e nessuno dei partecipanti aveva alcuna malattia mentale o fisica. Tutti i partecipanti avevano una visione normale o una visione corretta superiore a 1.0 e punteggi di intelligenza > 110 sul test Raven. Prima dell’esperimento, lo sperimentatore ha spiegato la procedura e punti di attenzione ai partecipanti per assicurarsi che avessero familiarità con il metodo e la procedura. Nelle settimane 2 prima dell’esperimento, i partecipanti dormivano regolarmente per 7-9 h al giorno, senza fumare, bere caffè, bere alcolici o consumare qualsiasi farmaco per 2 giorni prima dell’esperimento. Prima dell’esperimento, tutti i partecipanti hanno fornito il consenso informato scritto. Lo schema sperimentale è stato approvato dal Comitato Etico della Quarta Università medica militare e dell’Università di Beihang.
Progettazione sperimentale
Tre tipi di attività di memoria di lavoro sono stati presentati a tutti i partecipanti. Erano attività di memoria di lavoro con pronuncia a due posteriori (vedi Figura 1), attività di memoria di lavoro spaziale a due posteriori (vedi Figura 2) e attività di memoria di lavoro con oggetto a due posteriori (vedi Figura 3). I materiali di stimolazione dei compiti erano 15 lettere inglesi senza maiuscole e minuscole che escludevano quelle con lettere simili, come L/l, M/m; piccoli quadrati neri; e 12 figure geometriche, rispettivamente. Tutti i materiali sono stati mostrati in colore nero su uno sfondo bianco, con un angolo visivo approssimativo di 1,5° × 1,5° (larghezza: 2,0 cm, altezza: 2,0 cm) sotteso. 122 sentieri sono stati compresi in ogni attività e, in ogni percorso, lo stimolo target è stato presentato per 400 ms due percorsi dopo la presentazione dello stimolo obiettivo, con il tempo di asincronia di inizio dello stimolo 1,600 ms (SOA) che è stato contrassegnato da un bianco “+.”Ai partecipanti è stato chiesto di fare clic con il pulsante sinistro del mouse quando lo stimolo obiettivo e l’obiettivo erano gli stessi (“corrispondenza”), mentre fare clic con il pulsante destro del mouse quando non lo erano (“mancata corrispondenza”). La condizione corrispondente o meno è stata presentata in un ordine pseudorandom con un rapporto 1: 1.
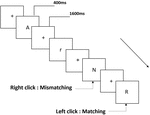
Figura 1. Diagramma schematico del compito di memoria di lavoro pronuncia.
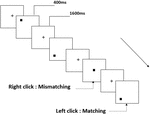
Figura 2. Diagramma schematico del compito di memoria di lavoro spaziale.
Figura 3. Diagramma schematico del compito di memoria di lavoro dell’oggetto.
Procedure sperimentali
Prima dell’esperimento, i partecipanti sono stati istruiti sul compito sperimentale. Sono stati informati di praticare i tre tipi di attività di memoria di lavoro fino a raggiungere un tasso di precisione del 90%. I partecipanti hanno visitato il laboratorio il giorno prima dell’esperimento e hanno dormito in laboratorio quella notte. I due partecipanti partner hanno eseguito gli esperimenti contemporaneamente. Tre tipi di attività di memoria di lavoro sono state eseguite dalle 7: 30 alle 8: 30 del mattino successivo con registrazione simultanea dell’elettroencefalogramma (EEG) (linea di base). La seconda registrazione EEG (36 h-TSD) è stata condotta dopo un periodo di 36 ore durante il quale ai partecipanti non è stato permesso di dormire. Durante l’intero periodo dell’esperimento, l’inibizione centrale e i farmaci stimolanti sono stati vietati. I partecipanti sono stati accompagnati, osservati e ricordati dal personale infermieristico per tenerli svegli durante tutta la sessione di TSD.
Registrazioni EEG
Un EEG continuo del cuoio capelluto è stato registrato utilizzando tappi di elettrodi posti in 64 posizioni utilizzando il sistema 10-20 con un amplificatore SynAmps2. I mastoidi bilaterali (A1 e A2) sono stati usati come riferimento e la fronte è stata usata come terreno. Gli EEG sono stati registrati a 1.000 Hz e l’impedenza di tutti i canali è stata mantenuta al di sotto di 5 kΩ. Quattro elettrodi aggiuntivi sono stati posizionati sopra e sotto gli occhi destro e sinistro per registrare un elettroculogramma bipolare verticale e orizzontale.
Analisi dei dati di esperimenti comportamentali
A causa di errori tecnici, due casi sono stati cancellati mentre altri 14 casi sono stati inclusi nella seguente analisi statistica. I dati comportamentali includevano il tempo di reazione medio, la velocità corretta e il numero corretto per unità di tempo. I dati comportamentali negli stati di base e 36 h-TSD sono stati registrati per l’analisi. Le analisi sono state eseguite da IBM SPSS (V22.2), dove è stata avviata l’analisi post-hoc di Bonferroni con il metodo ANOVA (reiterated measures analysis of variance) con Greenhouse-Geisser. I risultati statistici sono stati presentati come media e deviazione standard (DS).
Analisi dei dati EEG
Il programma Scan 4.3 è stato utilizzato per analizzare i dati EEG, in cui gli artefatti EEG del movimento oculare sono stati corretti con il metodo di riduzione degli artefatti oculari. Epoche che vanno da -100 a 800 ms dei dati EEG continui sono stati estratti e filtrati da un filtro passa banda da 0,5 a 30 Hz con la pendenza di frequenza di 24 dB / oct. Le prove in cui la tensione superava ± 100 µV sono state respinte e la linea di base è stata corretta a un’ampiezza media di 100 ms. I componenti EEG sono stati mediati e calcolati con solo le risposte corrette. I componenti ERP P2 (100-250 ms), N2 (150-350 ms) e P3 (250-450 ms) degli studi di stimolo sono stati identificati e quantificati. Le ampiezze e le latenze di picco grand-medie dei componenti N2 e P3 sono state calcolate separatamente a F3, Fz, F4, C3, Cz, C4, P3, Pz e P4, e la componente P2 è stata calcolata a F3, Fz, F4, C3, Cz e C4 (Casement et al., 2006; Verweij et al., 2014).
Misure ripetute ANOVA è stato impiegato per tutti i risultati ERP. Gli effetti principali e le interazioni tra gli stati di sonno (baseline e 36 h-TSD), attività (pronuncia memoria di lavoro, memoria di lavoro spaziale, e oggetto di memoria di lavoro), regioni (frontali, centrale e parietale; P2 componente è stato analizzato solo sul frontale, centrale e regioni), siti (di sinistra, centro e destra) sono stati analizzati statisticamente impiego di misure ripetute ANOVA, che comprendeva Serra-Geisser correzioni per non sfericità e di Bonferroni post-hoc test.
Risultati
Prestazioni comportamentali
I risultati degli esperimenti comportamentali sono mostrati nella Tabella 1. Il tempo di reazione medio è stato più lungo nello stato di 36 h-TSD rispetto al basale, con una tendenza all’aumento ma senza differenze significative. ANOVA ha rivelato che il tasso corretto del compito era significativamente diverso tra la linea di base e 36 h-TSD . Il numero corretto per unità di tempo ha mostrato un significativo effetto principale del tempo durante 36 h-TSD .

Tabella 1. Dati sulle prestazioni (media ± SD) sull’attività 2-back al basale e dopo 36 h-TSD.
Ampiezza
Rispetto al basale, è stata osservata una diminuzione significativa dell’ampiezza di P3 e un aumento significativo dell’ampiezza di P2 dopo TSD. Sebbene l’ampiezza N2 sia diminuita dopo 36 h di TSD, la differenza non ha raggiunto la significatività statistica (Tabella 2).

Tabella 2. Grand-ampiezza di picco media dei componenti P2, N2 e P3 nella condizione di risposta corretta su più siti di elettrodi al basale e dopo 36 h-TSD.
Significativi effetti principali di regioni e siti sull’ampiezza P2 sono stati trovati nella condizione TSD. Durante il TSD, l’ampiezza massima di P2 è apparsa nella regione frontale (Figura 4). Inoltre, le differenze nelle ampiezze P2 nelle diverse regioni (frontale vs centrale) erano più significative negli elettrodi bilaterali (a sinistra: P = 0,001; a destra: P = 0,000) rispetto agli elettrodi centrali (Figura 4). Un significativo effetto principale della regione e del sito su N2 ha rivelato che l’ampiezza N2 era più negativa nel frontale che nella regione centrale (P = 0,008, Figura 4) ed era più piccola a destra che a sinistra (P = 0,011, Figure 5A2,B2). È stato osservato un effetto principale del sito sull’ampiezza P3 . L’ampiezza di P3 era più positiva nel mezzo che sul lato sinistro (P = 0,009, Figura 4). Un effetto significativo di interazione tra tempo e regione è stato osservato per l’ampiezza P3 . Durante il TSD, la riduzione dell’ampiezza P3 era più significativa nelle regioni frontali e centrali rispetto alla regione parietale (P = 0,005; P = 0,003) (Figura 5C3). Nessun altro effetto principale o effetto di interazione ha raggiunto la significatività statistica.
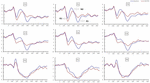
Figura 4. Ampiezza ERP al basale e 36 h-TSD per la condizione di risposta corretta per l’attività di memoria di lavoro. I canali sono ordinati da sinistra a destra e dall’alto verso il basso come segue: F3, Fz, F4, C3, Cz, C4, P3, Pz e P4. Rispetto al basale, le latenze dei componenti N2-P3 sono state prolungate e le ampiezze di N2-P3 sono diminuite dopo 36 h-TSD.
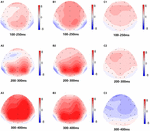
Figura 5. Mappa topografica della risposta corretta nell’attività di memoria di lavoro in diverse condizioni di sonno (A1–C3). (A1) P2, 100-250 ms, al basale. (A2) N2, 200-300 ms, al basale. (A3) P3, 300-400 ms, al basale. (B1) P2, 100-250 ms, a 36 h-TSD. (B2) N2, 200-300 ms, per 36 h-TSD. (B3) P3, 300-400 ms, per 36 h-TSD. (C1) P2, 100-250 ms, 36 h-TSD con la linea di base sottratta. (C2) N2, 200-300 ms, 36 h-TSD con linea di base sottratta. (C3) P3, 300-400 ms, 36 h-TSD con la linea di base sottratta.
Latenza
Le latenze di N2 e P2 sono state significativamente prolungate dopo TSD. Sebbene la latenza P3 sia stata prolungata dopo 36 ore di TSD, la differenza non ha raggiunto la significatività statistica (Tabella 3).

Tabella 3. Grand-latenza di picco media dei componenti P2, N2 e P3 nella condizione di risposta corretta su più siti di elettrodi al basale e dopo 36 h-TSD.
L’effetto principale significativo della regione su N2 e P3 ha rivelato che la latenza dei componenti N2-P3 era più breve nella regione parietale rispetto alla regione frontale (P = 0,002; P = 0,000) e alla regione centrale (P = 0,000; P = 0,000) (Figura 4). La latenza dell’onda P3 era significativamente più lunga sul lato sinistro che sul lato destro (Figura 4).
Nessun altro effetto principale o effetto di interazione ha raggiunto la significatività statistica.
Le ampiezze e le latenze N2, P2 e P3 che sono state suscitate nei nove siti degli elettrodi sono presentate in Figura 4. La mappa topografica della risposta corretta nell’attività di memoria di lavoro in diverse condizioni di sonno (linea di base, 36 h-TSD e la differenza tra le due condizioni) è presentata in Figura 5.
Discussione
In questo studio, abbiamo riportato le influenze della privazione del sonno 36 h sulla memoria di lavoro, combinando dati comportamentali in due stati di sonno (basale e 36 h-TSD) con registrazioni EEG contemporanee. L’analisi dei risultati ha indicato che i cambiamenti nei dati comportamentali in accordo con la memoria di lavoro compromessa dopo 36 h TSD: un aumento del tempo di reazione medio dei compiti cognitivi e una diminuzione della precisione.
La privazione del sonno ha compromesso il controllo individuale delle risorse attentive. Sebbene gli individui abbiano cercato di mantenere la veglia e le prestazioni lavorative, incluso il tempo di reazione e la velocità corretta, durante la privazione del sonno, la capacità di elaborazione delle informazioni della loro memoria di lavoro era ancora influenzata a causa della diminuzione della velocità di elaborazione delle informazioni (Casement et al., 2006; Wiggins et al., 2018). In questo studio, le onde N2 e P3 relative alla memoria di lavoro sono state misurate per mostrare un aumento della latenza e una diminuzione dell’ampiezza dopo la privazione del sonno rispetto alle letture di base. Gli studi hanno dimostrato che la privazione del sonno porta ad un continuo declino dell’attenzione e il fenomeno della diminuzione dell’ampiezza P3 indica che il controllo top-down della cognizione degli individui crolla gradualmente. La privazione del sonno ha un effetto più negativo sulle funzioni cognitive, specialmente quelle che dipendono da cognizioni o cognizioni mentali (Kusztor et al., 2019).
Il componente P3 riflette la distribuzione delle risorse di attenzione e la latenza di P3 è ampiamente vista come la finestra temporale per la categorizzazione e la valutazione dello stimolo. La diminuzione dell’ampiezza dell’onda P3 ha anche confermato che il processo decisionale nella risposta di corrispondenza dopo TSD era stato danneggiato in una certa misura (Gosselin et al., 2005). Gli studi hanno suggerito che la privazione del sonno può influenzare la fase di elaborazione delle informazioni della memoria di lavoro. In questo studio, gli indicatori di performance hanno anche supportato la conclusione che il tempo di risposta allo stimolo target è stato aumentato e che la latenza dell’onda P3 è stata prolungata (Cote et al., 2008). È stato ipotizzato che l’effetto della privazione del sonno sui componenti P3 potrebbe verificarsi anche a causa della mancata risposta all’alterazione delle informazioni, che è coerente con le precedenti conclusioni secondo cui i componenti P3 sono correlati all’aggiornamento del contenuto della memoria di lavoro (Donchin e Fabiani, 1991).
Studi precedenti hanno considerato il componente N2 come un indice elettrofisiologico che riflette la capacità dell’individuo di sopprimere la risposta (Kreusch et al., 2014). Dopo la privazione del sonno, la latenza prolungata del componente NoGo-N2 indica che la capacità dell’individuo di sopprimere la risposta è compromessa (Jin et al., 2015). La diminuzione dell’ampiezza e la latenza prolungata del componente N2 relativo alla memoria di lavoro della pronuncia dopo la privazione del sonno rivela che la privazione del sonno compromette l’elaborazione delle informazioni sulla memoria di lavoro della pronuncia (Zhang et al., 2019). La componente N2 è generalmente pensata per riflettere l’attenzione selettiva del cervello e l’elaborazione di stimoli o segnali emotivi (Schacht et al., 2008) ed è una componente endogena correlata allo stato mentale, all’attenzione e al grado di attenzione di un individuo. In questo studio, abbiamo scoperto che la latenza del componente N2 era significativamente prolungata, ma l’ampiezza mostrava solo una tendenza al ribasso. Secondo studi precedenti, la latenza prolungata di N2 rifletteva un aumento del tempo di risposta dopo la restrizione del sonno (Zhang et al., 2014). Tuttavia, la scoperta che l’ampiezza N2 non era significativamente alterata potrebbe essere stata dovuta a risposte compensative cerebrali (Drummond e Brown, 2001). Nel caso di risorse cognitive limitate, c’era un meccanismo di compensazione per ripristinare la funzione cognitiva compromessa (Jin et al., 2015).
Secondo la topografia del cuoio capelluto, i cambiamenti nei componenti N2-P3 relativi alla privazione del sonno sono più evidenti nell’area frontale. Il controllo frontoparietale (FPC) svolge un ruolo importante nel controllo cognitivo. Gli studi hanno dimostrato che l’FPC può bypassare il controllo cognitivo dall’alto verso il basso, consentendo agli individui di concentrarsi sulle informazioni relative al bersaglio sopprimendo le informazioni che non sono correlate al bersaglio (Smallwood et al., 2011; Wen et al., 2013). L’FPC è importante per la conservazione delle informazioni e l’elaborazione delle informazioni nella memoria di lavoro e il grado di attivazione dell’FPC dopo la privazione del sonno è stato ridotto rispetto a un normale gruppo di sonno (Ma et al., 2014). Sebbene i risultati dell’EEG non riflettessero i cambiamenti in specifiche regioni del cervello in dettaglio, rifletteva intuitivamente l’effetto del TSD sulla conservazione e l’elaborazione delle informazioni sulla memoria di lavoro.
Sebbene l’esatto processo cognitivo alla base della componente P2 sia ancora ampiamente dibattuto, come definizione ampia, la componente P2 riflette il processo di attenzione e elaborazione visiva ed è generalmente considerata correlata all’attenzione selettiva e alla memoria di lavoro, riflettendo il giudizio precoce del processo percettivo (Saito et al., 2001). In questo studio, abbiamo trovato un aumento significativo dell’ampiezza dell’onda P2 dopo la privazione del sonno. Gli studi hanno riferito che le onde P2 che potrebbero essere una parte del sistema di corrispondenza cognitiva precoce per l’elaborazione dei messaggi e possono confrontare gli input sensoriali con la memoria memorizzata (Freunberger et al., 2007) sono sensibili alle alterazioni dell’attenzione alla missione e alle richieste di memoria di lavoro (Smith et al., 2002). La compensazione funzionale è una delle funzioni uniche del cervello umano e un fattore importante per mantenere la funzione cognitiva. L’eccessiva attivazione della corteccia prefrontale dorsolaterale (DLPFC) dopo la privazione del sonno indica che, con la diminuzione delle risorse cerebrali, la DLPFC sembra avere una funzione compensatoria (Drummond et al., 2004; Choo et al., 2005). Pertanto, ipotizziamo che l’aumento significativo dell’ampiezza P2 osservato in questo studio possa essere dovuto alla compensazione funzionale in cui gli individui sembrano mantenere la normale funzione cognitiva dopo la privazione del sonno. Sebbene un gran numero di studi abbia utilizzato la tecnologia ERP per esplorare l’effetto della privazione del sonno sulle funzioni cognitive, i primi componenti come N1 e P2 non sono stati studiati sistematicamente e i risultati sono incoerenti (Evans e Federmeier, 2007; Wiggins et al., 2018; Zhang et al., 2019). Ci sono poche ricerche che esplorano il cambiamento della componente P2 durante la privazione del sonno (Mograss et al., 2009). Pertanto, gli effetti della privazione del sonno sui primi componenti di ERP, come P2, devono ancora essere ulteriormente studiati ed esplorati.
In questo esperimento, abbiamo usato il modello 2-back per progettare compiti di pronuncia, spazio e memoria di lavoro degli oggetti e abbiamo esaminato la compromissione della memoria di lavoro dopo 36 ore di TSD. Rispetto agli studi precedenti che si concentravano solo sugli effetti della privazione del sonno su un tipo specifico di informazioni, come la pronuncia della memoria di lavoro o una funzione cognitiva specifica, come l’inibizione della risposta, abbiamo considerato il contenuto del modello di memoria di lavoro e analizzato in modo completo gli effetti della privazione del sonno sulla memoria di lavoro.
Tuttavia, lo studio presenta alcune limitazioni. Innanzitutto, abbiamo utilizzato solo l’attività 2-back e non siamo riusciti a confrontare le prestazioni dei partecipanti nelle attività di memoria di lavoro di diverse difficoltà. Pertanto, ci sono limitazioni nello spiegare e dedurre i cambiamenti nel carico di lavoro. In secondo luogo, nello studio sono stati utilizzati solo volontari di sesso maschile e le conclusioni devono essere valutate quando le si estende alle volontarie di sesso femminile. A causa del numero limitato di partecipanti, abbiamo scoperto solo che l’ampiezza dell’onda N2 aveva una tendenza al ribasso e che la latenza dell’onda P3 aveva una tendenza prolungata. Risultati stabili potrebbero essere ottenuti dopo aver aumentato il numero di partecipanti. In terzo luogo, la combinazione della nostra procedura con fMRI per la memoria di lavoro può facilitare un’ulteriore interpretazione dei risultati. Studi precedenti hanno dimostrato che i bioritmi circadiani influenzano le prestazioni comportamentali e ci sono differenze individuali (Montplaisir, 1981; Lavie, 2001). Non abbiamo registrato i dati EEG nello stesso momento in questo esperimento, quindi l’influenza dei bioritmi circadiani sui risultati del test non può essere completamente esclusa.
Questa ricerca ha dimostrato che la capacità di memoria di lavoro era compromessa dopo TSD e che questo danno non era associato al contenuto di stimolo della memoria di lavoro. La mancanza di sonno ha ridotto la qualità delle informazioni memorizzate nella memoria, che potrebbe verificarsi con il processo degenerativo di attenzione (Ratcliff e Van Dongen, 2018). Questo studio fornisce prove di elettrofisiologia per comprendere il meccanismo sotto la memoria di lavoro compromessa dopo la privazione del sonno. È necessario prestare attenzione agli effetti negativi della compromissione della memoria di lavoro causata dalla privazione del sonno e esplorare interventi efficaci per tale danno.
Dichiarazione di disponibilità dei dati
I set di dati generati per questo studio sono disponibili su richiesta all’autore corrispondente.
Dichiarazione etica
Gli studi che hanno coinvolto partecipanti umani sono stati esaminati e approvati dalla Quarta Università medica militare Beihang University. I pazienti / partecipanti hanno fornito il loro consenso informato scritto a partecipare a questo studio.
Contributi dell’autore
YS ha progettato gli esperimenti. ZP ha prodotto i risultati e ha scritto il manoscritto. CD e LZ hanno analizzato e interpretato i dati. JT e YS hanno eseguito gli esperimenti, familiarizzato con i dati e con i garanti di questo studio. YB, LZ e JT hanno contribuito a partecipare alla raccolta dei dati e alla revisione della letteratura. Tutti gli autori elencati hanno letto e approvato il manoscritto finale.
Finanziamento
Questa ricerca è stata sostenuta dalla National Science Foundation of Winter Olympics Technology Plan of China sotto Grant Nos. 2019YFF0301600 e HJ20191A020135.
Conflitto di interessi
Gli autori dichiarano che la ricerca è stata condotta in assenza di rapporti commerciali o finanziari che potrebbero essere interpretati come un potenziale conflitto di interessi.
Anderson, C., e Platten, CR (2011). La privazione del sonno riduce l’inibizione e migliora l’impulsività agli stimoli negativi. Comportati bene. Cervello Res. 217: 466. doi: 10.1016 / j.bbr.2010.09.020
PubMed Abstract | CrossRef Full Text/Google Scholar
Baddeley, A. (2000). Il buffer episodico: un nuovo componente della memoria di lavoro? Tendenze Cogn. Sic. 4:417. il nostro sito utilizza cookie tecnici e di terze parti per migliorare la tua esperienza di navigazione e per migliorare la tua esperienza di navigazione. Attenzione e memoria di lavoro negli anestesisti residenti dopo il servizio notturno: effetti di gruppo e individuali. Occup. Ambiente. Med. 61, 167–170. doi: 10.1136 / oem.2002.006197
PubMed Abstract | CrossRef Full Text/Google Scholar
Casement, MD, Broussard, JL, Mullington, JM, and Press, DZ (2006). Il contributo del sonno ai miglioramenti nella velocità di scansione della memoria di lavoro: uno studio sulla restrizione prolungata del sonno. Biol. Psicol. 72:212. doi: 10.1016 / j. biopsycho.2005.11.002
PubMed Abstract | CrossRef Full Text/Google Scholar
Chee, M. W. L., e Chuah, L. Y. M. (2008). Neuroimaging funzionale intuizioni su come il sonno e la privazione del sonno influenzano la memoria e la cognizione. Curr. Opin. Neurolo. 21, 417–423. doi: 10.1097 / OMD.0b013e3283052cf7
PubMed Abstract | CrossRef Full Text/Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, FS, e Chee, M. W. L. (2005). Dissociazione delle regioni corticali modulata sia dal carico di memoria di lavoro che dalla privazione del sonno e dalla sola privazione del sonno. Neuroimage 25, 579-587. doi: 10.1016 / j.neuroimage.2004.11.029
PubMed Abstract | CrossRef Full Text/Google Scholar
Chuah, YML, Venkatraman, V., Dinges, DF, e Chee, MWL (2006). La base neurale della variabilità interindividuale nell’efficienza inibitoria dopo la privazione del sonno. J. Neurosci. 26, 7156–7162. doi: 10.1523 / jneurosci.0906-06. 2006
CrossRef Full Text/Google Scholar
Cote, K. A., Milner, CE, Osip, SL, Baker, ML, and Cuthbert, B. P. (2008). Eccitazione fisiologica e attenzione durante una settimana di restrizione continua del sonno. Physiol. Comportati bene. 95, 353–364. doi: 10.1016 / j.physbeh.2008.06.016
PubMed Abstract | CrossRef Full Text/Google Scholar
Cote, K. A., Mondloch, CJ, Sergeeva, V., Taylor, M., e Semplonius, T. (2014). Impatto della privazione totale del sonno sull’elaborazione neurale comportamentale di volti emotivamente espressivi. Scad. Brain Res. 232, 1429-1442. doi: 10.1007 / s00221-013-3780-1
PubMed Abstract | CrossRef Full Text/Google Scholar
Donchin, E., e Fabiani, M. (1991). L’uso dei potenziali cerebrali correlati agli eventi nello studio della memoria: p300 è una misura del carattere distintivo degli eventi? Psicol. Facul. Pubblico. 7, 58–77. doi: 10.1111 / sltb.12055
PubMed Abstract | CrossRef Full Text/Google Scholar
Drummond, S. P., Brown, G. G., Salamat, J. S., and Gillin, J. C. (2004). L’aumento della difficoltà del compito facilita la risposta compensatoria cerebrale alla privazione totale del sonno. Dormire 27, 445-451. doi: 10.1093/sleep/27.3.445
CrossRef Full Text/Google Scholar
Drummond, S. P. A., e Brown, G. G. (2001). Gli effetti della privazione totale del sonno sulle risposte cerebrali alle prestazioni cognitive. Neuropsicofarmacologia 25, S68-S73. doi: 10.1016/S0893-133X (01)00325-6
CrossRef Full Text/Google Scholar
Evans, KM, e Federmeier, KD (2007). La memoria a destra e la memoria a sinistra: i potenziali legati agli eventi rivelano asimmetrie emisferiche nella codifica e nella conservazione delle informazioni verbali. Neuropsicologia 45, 1777-1790. doi: 10.1016 / j.neuropsicologia.2006.12.014
PubMed Abstract | CrossRef Full Text/Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M., and Höller, Y. (2007). Il componente p2 visivo è correlato al blocco di fase theta. Neurosci. Lett. 426, 181–186. doi: 10.1016 / j.neulet.2007.08.062
PubMed Abstract | CrossRef Full Text/Google Scholar
Géraldine, R., Desgranges, B., Foret, J., and Eustache, F. (2005). Le relazioni tra i sistemi di memoria e le fasi del sonno. J. Sleep Res. 14, 123-140. doi: 10.1111 / j. 1365-2869. 2005.00450.per questo motivo, è stato creato un sito web dedicato alla creazione di siti web e di siti web per la creazione di siti web e di siti web per la gestione di siti web e siti web. Privazione totale del sonno ed elaborazione della novità: implicazioni per il funzionamento del lobo frontale. Clin. Neurofisiolo. 116, 211–222. doi: 10.1016 / j.clinph.2004.07.033
PubMed Abstract | CrossRef Full Text/Google Scholar
Harrison, Y., e Horne, J. (1998). La perdita di sonno compromette le attività linguistiche brevi e nuove con un focus prefrontale. J. Sleep Res. 7, 95-100. doi: 10.1046 / j. 1365-2869. 1998.00104.per maggiori informazioni, consultare il sito:
PubMed Abstract/CrossRef Full Text/Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., and Chamari, K. (2013). L’impatto della privazione parziale del sonno sulle variazioni diurne delle prestazioni cognitive nei soggetti allenati. Proc. Soc. Comportati bene. Sic. 82, 392–396. doi: 10.1016 / j. sbspro.2013.06.281
CrossRef Full Text/Google Scholar
Jennings, J. R., Monk, T. H., and Molen, M. W. V. D. (2003). La privazione del sonno influenza alcuni ma non tutti i processi di attenzione di supervisione. Psicol. Sic. 14, 473–479. doi: 10.1111 | 1467-9280.02456
PubMed Abstract/CrossRef Full Text/Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L., e Lei, Y. (2015). Il sonno di recupero inverte l’inibizione della risposta alterata a causa della restrizione del sonno: evidenza da uno studio sui potenziali correlati agli eventi visivi. PLoS Uno 10:e0142361. doi: 10.1371 / giornale.pone.0142361
PubMed Abstract | CrossRef Full Text/Google Scholar
Jones, K., e Harrison, Y. (2001). Funzione del lobo frontale, perdita di sonno e sonno frammentato. Dormire Med. Apoc 5:475. doi: 10.1053 / smrv.2001.0203
PubMed Abstract | CrossRef Full Text/Google Scholar
Koslowsky, M., and Babkoff, H. (1992). Meta-analisi della relazione tra privazione totale del sonno e prestazioni. Cronobiolo. Int. 9, 132–136. doi: 10.3109/07420529209064524
PubMed Abstract | CrossRef Full Text/Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A., and Hansenne, M. (2014). Abuso di alcol e componenti erp in attività go/no-go utilizzando stimoli correlati all’alcol: impatto dell’evitamento dell’alcol. Int. J. Psicofisiolo. 94, 92–99. doi: 10.1016 / j. ijpsycho.2014.08.001
PubMed Abstract | CrossRef Full Text/Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, AS, Storm, J. F., and Huster, R. J. (2019). La privazione del sonno influisce in modo differenziato sui sottocomponenti del controllo cognitivo. Dormire 42: zsz016. doi: 10.1093/sleep/zsz016
PubMed Abstract / CrossRef Full Text/Google Scholar
Lavie, P. (2001). Sonno-veglia come ritmo biologico. Annu. Rev. Psicol. 52, 277–303. doi: 10.1146 / annurev.psych.52.1.277
CrossRef Full Text/Google Scholar
Lei, Y., Shao, Y., Wang, L., Zhai, T., e Zou, F. (2015). L’accoppiamento di rete cerebrale su larga scala predice gli effetti di privazione totale del sonno sulla capacità cognitiva. PLoS Uno 10:e0133959. doi: 10.1371 / giornale.pone.0133959
PubMed Abstract | CrossRef Full Text/Google Scholar
Li, C., Huang, D., Qi, J., Chang, H., e Meng, Q. (2016). I deficit di memoria a breve termine sono correlati con alterazioni della connettività funzionale ippocampale-talamica a seguito di restrizione acuta del sonno. L’imaging cerebrale si comporta. 11, 954–963. doi: 10.1007 / s11682-016-9570-1
PubMed Abstract | CrossRef Full Text/Google Scholar
Ma, N., Dinges, D. F., Basner, M., and Rao, H. (2014). Come acuta perdita di sonno totale colpisce il cervello che frequentano: una meta-analisi di studi di neuroimaging. Dormire 38, 233. doi: 10.5665 / dormire.4404
PubMed Abstract | CrossRef Full Text/Google Scholar
Miyake, A., e Shah, P. (1999). Modelli di memoria di lavoro. Meccanismi di Manutenzione Attiva e Controllo Esecutivo. Cambridge: Cambridge University Press.
Google Scholar
Mograss, M. A., Guillem, F., Brazzini-Poisson, V., and Godbout, R. (2009). Gli effetti della privazione totale del sonno sui processi di memoria di riconoscimento: uno studio del potenziale correlato agli eventi. Neurobiolo. Imparare. Mem. 91, 343–352. doi: 10.1016 / j. nlm.2009.01.008
PubMed Abstract | CrossRef Full Text/Google Scholar
Montplaisir, J. (1981). Depressione e ritmo biologico: effetti terapeutici della privazione del sonno. L’union Médicale du Canada 110, 272-276. doi: 10.1620 / tjem.133.481
PubMed Abstract | CrossRef Full Text/Google Scholar
Morris, A. M., So, Y., Lee, K. A., Lash, A. A., e Becker, E. V. (1992). Il potenziale legato all’evento p300. gli effetti della privazione del sonno. J. Occupat. Med. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F., e Hobson, J. A. (2002). La corteccia prefrontale nel sonno. Tendenze Cogn. Sic. 6, 475–481. doi: 10.1016/S1364-6613(02)01992-7
CrossRef Full Text/Google Scholar
Ning, M., Dinges, DF, Mathias, B. e Hengyi, R. (2014). Come acuta perdita di sonno totale colpisce il cervello che frequentano: una meta-analisi di studi di neuroimaging. Sonno 38, 233-240. doi: 10.5665 / dormire.4404
PubMed Abstract | CrossRef Full Text/Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, A. R., and Bullmore, E. (2005). Paradigma della memoria di lavoro N-back: una meta-analisi di studi di neuroimaging funzionale normativo. Ronzio. Cervello Mapp. 25, 46–59. doi: 10.1002 / hbm.Nel 2013, il progetto è stato completato con la pubblicazione di un testo completo di CrossRef/Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S., e Kumar, S. (2010). I correlati elettrofisiologici della cognizione migliorano con il pisolino durante la privazione del sonno. EUR. J. Appl. Physiol. 108, 549–556. doi: 10.1007 / s00421-009-1222-3
PubMed Abstract | CrossRef Full Text/Google Scholar
Ratcliff, R., and Van Dongen, H. (2018). Gli effetti della privazione del sonno sulla voce e la memoria di riconoscimento associativo. J. Scad. Psicol. 44, 193–208. doi: 10.1037/xlm0000452
PubMed Abstract / CrossRef Full Text/Google Scholar
Raymond, CA (1988). Spostando il lavoro, i cicli del sonno sono sulla strada per diventare un altro problema di salute pubblica. JAMA J. Am. Med. Assoc. 259, 2958–2959. doi: 10.1001 / jama.In questo modo, il sistema di gestione delle risorse umane è in grado di garantire la sicurezza e la sicurezza di tutti gli utenti. Potenziale visivo correlato agli eventi nella demenza lieve di tipo alzheimer. Psichiatria Clin. Neurosci. 55, 365–371. doi: 10.1046 / j. 1440-1819. 2001.00876.per ulteriori informazioni, visitare il sito:
PubMed Abstract | CrossRef Full Text/Google Scholar
Smallwood, J., Brown, K., Baird, B., e Schooler, JW (2011). Cooperazione tra la rete di modalità predefinita e la rete frontale-parietale nella produzione di un treno di pensiero interno. Cervello Res. 1428, 60-70. doi: 10.1016 / j. brainres.2011.03.072
PubMed Abstract | CrossRef Full Text/Google Scholar
Smith, M. E., Mcevoy, L. K., and Gevins, A. (2002). L’impatto della moderata perdita di sonno sui segnali neurofisiologici durante le prestazioni delle attività di memoria di lavoro. Dormire 25, 784-794. doi: 10.1093 / dormire/25.7.56
CrossRef Full Text/Google Scholar
Thomas, M. (2003). Base neurale di vigilanza e compromissione delle prestazioni cognitive durante la sonnolenza ii. effetti di 48 e 72 ore di privazione del sonno sulla veglia dell’attività cerebrale regionale umana. Thalam. Relat. Syst. 2, 199–229. doi: 10.1016/S1472-9288(03)00020-7
CrossRef Full Text/Google Scholar
Van Dongen, H. (2004). Il costo cumulativo della veglia aggiuntiva: doseresponse effetti sulle funzioni neurocomportamentali e fisiologia del sonno da privazione cronica del sonno e privazione totale del sonno. Dormire 26, 117-126. doi: 10.1016/0739-6260(84)90040-6il nostro sito utilizza cookie tecnici e di terze parti per migliorare la tua esperienza di navigazione. La privazione del sonno porta a una perdita di connettività funzionale nelle regioni cerebrali frontali. BMC Neurosci. 15:88. doi: 10.1186 | 1471-2202-15-88
PubMed Abstract / CrossRef Full Text/Google Scholar
Wen, X., Liu, Y., Yao, L., e Ding, M. (2013). Regolazione dall’alto verso il basso dell’attività della modalità predefinita nell’attenzione visiva spaziale. J. Neurosci. 33, 6444–6453. doi: 10.1523 / JNEUROSCI.Il nostro sito utilizza cookie tecnici e di terze parti per migliorare la tua esperienza di navigazione. la privazione del sonno 24-h compromette la modulazione attentiva precoce dell’elaborazione neurale: uno studio del potenziale cerebrale correlato agli eventi. Neurosci. Lett. 677, 32–36. doi: 10.1016 / j.neulet.2018.04.022
PubMed Abstract | CrossRef Full Text/Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y. e Zhou, Q. (2019). Diminuzione delle informazioni sostituzione della memoria di lavoro dopo la privazione del sonno: evidenza da uno studio potenziale correlato agli eventi. Anteriore. Neurosci. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef Full Text/Google Scholar
Zhang, Q., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z., et al. (2014). Uno studio erp visivo di inibizione dell’impulso a seguito di un pisolino indotto da zaleplon dopo la privazione del sonno. PLoS Uno 9: e95653. doi: 10.1371 / giornale.pone.0095653
PubMed Abstract / CrossRef Full Text / Google Scholar
Leave a Reply