Bookshelf
Nonostante la sua posizione periferica, la retina o la porzione neurale dell’occhio, è in realtà parte del sistema nervoso centrale. Durante lo sviluppo, la retina si forma come un outpocketing del diencefalo, chiamato vescicola ottica, che subisce l’invaginazione per formare la coppa ottica (Figura 11.3; vedi anche Capitolo 22). La parete interna della tazza ottica dà origine alla retina, mentre la parete esterna dà origine all’epitelio pigmentato. Questo epitelio è una struttura melanincontaining che riduce il backscattering di luce che entra nell’occhio; inoltre svolge un ruolo critico nel mantenimento dei fotorecettori, rinnovando i fotopigmenti e fagocitando i dischi del fotorecettore, di cui il giro d’affari ad un tasso elevato è essenziale alla visione.

Figura 11.3
Sviluppo dell’occhio umano. (A) La retina si sviluppa come un outpocketing dal tubo neurale, chiamato vescicola ottica. (B) La vescicola ottica invagina per formare la coppa ottica. (C, D) La parete interna della coppa ottica diventa la retina neurale, mentre il (più…)
Coerentemente con il suo status di parte a pieno titolo del sistema nervoso centrale, la retina comprende circuiti neurali complessi che convertono l’attività elettrica graduata dei fotorecettori in potenziali d’azione che viaggiano al cervello tramite assoni nel nervo ottico. Anche se ha gli stessi tipi di elementi funzionali e neurotrasmettitori trovati in altre parti del sistema nervoso centrale, la retina comprende solo poche classi di neuroni, e questi sono disposti in un modo che è stato meno difficile da svelare rispetto ai circuiti in altre aree del cervello. Ci sono cinque tipi di neuroni nella retina: fotorecettori, cellule bipolari, cellule gangliari, cellule orizzontali e cellule amacrine. I corpi e i processi cellulari di questi neuroni sono impilati in cinque strati alternati, con i corpi cellulari situati negli strati nucleari interni, nucleari esterni e gangliari e i processi e i contatti sinaptici situati negli strati plessiformi interni ed esterni (Figura 11.4). Una catena diretta di tre neuroni – cellula fotorecettrice a cellula bipolare a cellula gangliare-è la principale via di flusso di informazioni dai fotorecettori al nervo ottico.
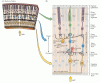
Figura 11.4
Struttura della retina. (A) Sezione della retina che mostra la disposizione complessiva degli strati retinici. (B) Schema dei circuiti di base della retina. Una catena di tre neuroni-fotorecettori, cellule bipolari e cellule gangliari-fornisce il più diretto (altro…)
Ci sono due tipi di elementi sensibili alla luce nella retina: bastoncelli e coni. Entrambi i tipi di fotorecettori hanno un segmento esterno che è composto da dischi membranosi che contengono fotopigmento e si trova adiacente allo strato epiteliale del pigmento, e un segmento interno che contiene il nucleo cellulare e dà origine a terminali sinaptici che contattano le cellule bipolari o orizzontali. L’assorbimento della luce da parte del fotopigmento nel segmento esterno dei fotorecettori avvia una cascata di eventi che cambia il potenziale di membrana del recettore e quindi la quantità di neurotrasmettitore rilasciato dalle sinapsi dei fotorecettori sulle cellule che contattano. Le sinapsi tra i terminali dei fotorecettori e le cellule bipolari (e le cellule orizzontali) si verificano nello strato plessiforme esterno; più specificamente, i corpi cellulari dei fotorecettori costituiscono lo strato nucleare esterno, mentre i corpi cellulari delle cellule bipolari si trovano nello strato nucleare interno. I brevi processi assonali delle cellule bipolari creano contatti sinaptici a loro volta sui processi dendritici delle cellule gangliari nello strato plessiforme interno. Gli assoni molto più grandi delle cellule gangliari formano il nervo ottico e portano informazioni sulla stimolazione retinica al resto del sistema nervoso centrale.
Gli altri due tipi di neuroni nella retina, le cellule orizzontali e le cellule amacrine, hanno i loro corpi cellulari nello strato nucleare interno e sono principalmente responsabili delle interazioni laterali all’interno della retina. Queste interazioni laterali tra i recettori, le cellule orizzontali e le cellule bipolari nello strato plessiforme esterno sono in gran parte responsabili della sensibilità del sistema visivo al contrasto della luminanza su un’ampia gamma di intensità luminose. I processi delle cellule amacrine, che si estendono lateralmente nello strato plessiforme interno, sono postsinaptici ai terminali delle cellule bipolari e presinaptici ai dendriti delle cellule gangliari (vedi Figura 11.4). I processi delle cellule orizzontali si ramificano nello strato plessiforme esterno. Diverse sottoclassi di cellule amacrine che danno contributi distinti alla funzione visiva. Una classe di cellule amacrine, ad esempio, svolge un ruolo importante nel trasformare le risposte persistenti delle cellule bipolari alla luce nelle brevi risposte transitorie esposte da alcuni tipi di cellule gangliari. Un altro tipo serve come passo obbligatorio nel percorso che trasmette informazioni dai fotorecettori dell’asta alle cellule gangliari retiniche. La varietà di sottotipi di cellule amacrine illustra la regola più generale che, sebbene ci siano solo cinque tipi di cellule retiniche di base, ci può essere una notevole diversità all’interno di un determinato tipo di cellule. Questa diversità è la base per percorsi che trasmettono diversi tipi di informazioni agli obiettivi centrali in modo parallelo.
A prima vista, la disposizione spaziale degli strati retinici sembra controintuitiva, poiché i raggi di luce devono passare attraverso gli elementi non sensibili alla luce della retina (e della vascolarizzazione retinica!) prima di raggiungere i segmenti esterni dei fotorecettori, dove i fotoni vengono assorbiti (vedi Figura 11.4). La ragione di questa curiosa caratteristica dell’organizzazione retinica risiede nella relazione speciale che esiste tra i segmenti esterni dei fotorecettori e l’epitelio pigmentato. I segmenti esterni contengono dischi membranosi che ospitano il fotopigmento sensibile alla luce e altre proteine coinvolte nel processo di trasduzione. Questi dischi si formano vicino al segmento interno del fotorecettore e si muovono verso la punta del segmento esterno, dove vengono versati. L’epitelio pigmentato svolge un ruolo essenziale nella rimozione dei dischi dei recettori consumati; questo non è un compito da poco, poiché tutti i dischi nei segmenti esterni vengono sostituiti ogni 12 giorni. Inoltre, l’epitelio pigmentato contiene il macchinario biochimico necessario per rigenerare le molecole di fotopigmento dopo che sono state esposte alla luce. Sono presumibilmente le esigenze del ciclo di vita del disco fotorecettore e del riciclaggio del fotopigmento che spiegano perché coni e bastoncelli si trovano nello strato più esterno piuttosto che nello strato più interno della retina. Le interruzioni nelle normali relazioni tra epitelio pigmentato e fotorecettori retinici, come quelle che si verificano nella retinite pigmentosa, hanno gravi conseguenze per la vista (Riquadro B).
Box B
Retinitis Pigmentosa.
Leave a Reply