Frontiers in Neuroscience
introduktion
med samfundets fremskridt og ændringer i arbejdsrytmen lider et stigende antal mennesker af søvnmangel. Søvnmangel skader ikke kun individets fysiske og mentale sundhed, men påvirker også arbejdsydelsen alvorligt og forårsager arbejdsfejl og endda ulykker. Derfor er forståelsen af mekanismen for søvnmangel, der påvirker kognitiv funktion, af stor betydning for effektivt at forhindre virkningerne af søvnmangel.
tidligere undersøgelser har afsløret, at søvnmangel kan forårsage en række ændringer i individets humør, kognitive evne, arbejdsindsats og immunfunktion (Choo et al., 2005). Manglen på søvn forstyrrer kroppens cirkulation og påvirker individers kognitive og følelsesmæssige evner (Raymond, 1988). Flere undersøgelser har afsløret, at søvnmangel forringer responsinhibering (Harrison and Horne, 1998; Musur et al., 2002; Jennings et al., 2003). For eksempel faldt individets evne til at undertrykke negative stimuli efter 36 timers søvnmangel (Chuah et al., 2006). Neuroimaging-undersøgelser har antydet, at søvnmangel reducerer en persons lave niveau af visuel behandlingsevne (Anderson and Platten, 2011; Ning et al., 2014). Derudover svækker søvnmangel hippocampus og kan påvirke hukommelsen ved at ødelægge synaptisk plasticitet (Cote et al., 2014). Thomas (2003) har indikeret, at mangel på søvn reducerede cerebral blodgennemstrømning og metabolisk hastighed i thalamus, præfrontal hjernebark og parietalbark (g., 2005). Jarraya og kolleger fandt ud af, at delvis søvnmangel signifikant påvirkede neuropsykologiske funktioner såsom verbal øjeblikkelig hukommelse, opmærksomhed og årvågenhed (Thomas, 2003). Desuden har nogle undersøgelser afsløret, at de kumulative virkninger af delvis søvnmangel alvorligt kan forringe kognitiv funktion og adfærd (Van Dongen, 2004; Scott et al., 2006; Jarraya et al., 2013).
arbejdshukommelse er et system, der bruges til at gemme og behandle information, og som er en kognitiv funktion med begrænset kapacitet (Bartel et al., 2004). Desuden kan de oplysninger, der er gemt i arbejdshukommelsessystemet, ændres fra korttidshukommelse til langtidshukommelse gennem genfortælling og andre hukommelsesmetoder. Arbejdshukommelse er overgangen mellem kortsigtede og langsigtede hukommelsessystemer, som er meget afgørende i menneskelig meddelelsesbehandling (Miyake og Shah, 1999). Det giver en midlertidig lagerplads og de ressourcer, der er nødvendige for at behandle information, såsom stemmeforståelse, ræsonnement og læring. Søvnmangel har vist sig at påvirke arbejdshukommelsen først.
tidligere undersøgelser har brugt N-back-arbejdshukommelsesparadigmet hos deltagere, der gennemgik søvnmangel og fandt, at mangel på søvn inducerer et fald i metabolisk aktivitet i hjernens regionale netværk, som hovedsageligt udføres informationsbehandling og reaktionshæmning (Baddeley, 2000; Jhang et al., 2019). Nedsat arbejdshukommelse efter søvnmangel er relateret til aktiveringen af standardnetværket i opgaver (Chee og Chuah, 2008), som kan være relateret til thalamusens vigtige rolle i kortikal opmærksomhed. For eksempel øgede søvnmangel forbindelsen mellem hippocampus, thalamus og standardnetværk, som ofte blev ledsaget af højere subjektiv døsighed og dårligere ydeevne af arbejdshukommelse (Lei et al., 2015; Li et al., 2016). Undersøgelser om søvnmangel identitet?ed, at øget latenstid og reduceret amplitude af P3-komponenten var forbundet med langvarig ædruelighed (Morris et al., 1992; Jones og Harrison, 2001; Panjvani et al., 2010). Faldet i P3-bølgen kan afspejle et fald i deltagernes opmærksomhed og en reduktion i skelnen mellem målstimuleringer (Koslovsky og Babkoff, 1992).
imidlertid har få undersøgelser leveret elektrofysiologisk bevis for nedsat arbejdshukommelse efter søvnmangel. N-back-opgaven betragtes som en almindelig metode til vurdering af arbejdshukommelse., 2005; Jaeggi et al., 2010). Jang et al. designet en to-back udtale arbejdshukommelsesopgave for at udforske den nedsatte besked alternativ til arbejdshukommelse under søvnmangel, men få undersøgelser har brugt forskellige typer arbejdshukommelsesopgaver i et enkelt eksperiment. I denne undersøgelse designede vi forskellige typer arbejdshukommelsesopgaver (udtale arbejdshukommelse, rumlig arbejdshukommelse og objektarbejdshukommelse) for at undersøge svækkelse af kognitiv funktion af TSD og registreret deltager EEG data på 2 tidspunkter (baseline og 36 h-TSD). Alle opgaverne vedtog et 2-back paradigme. Denne undersøgelse evaluerede ændringerne i N2-P3-bølgen relateret til arbejdshukommelse under TSD og analyserede de tidsmæssige egenskaber ved virkningerne af søvnmangel på arbejdshukommelsen. Vores resultater giver eksperimentelle beviser for virkningerne af søvnmangel på kognitiv funktion.
materialer og metoder
deltagere
seksten unge, sunde, højrehåndede mandlige studerende deltog i denne undersøgelse. Vi rekrutterede deltagere ved at annoncere på campus. Deltagerne havde alle gode søvnvaner (PSKI< 5). Alle deltagere var mellem 21 og 28 år med en gennemsnitlig alder på 23 år, og ingen af deltagerne havde nogen mental eller fysisk sygdom. Alle deltagere havde normalt syn eller korrigeret syn over 1,0 og intelligensresultater >110 på Raven-testen. Før eksperimentet forklarede eksperimentatoren proceduren og peger på deltagerne for at sikre, at de var fortrolige med metoden og proceduren. I de 2 uger før eksperimentet sov deltagerne regelmæssigt i 7-9 timer om dagen uden at ryge, drikke kaffe, drikke alkohol eller indtage medicin i 2 dage før eksperimentet. Før eksperimentet gav alle deltagerne skriftligt informeret samtykke. Den eksperimentelle ordning blev godkendt af det etiske udvalg for det fjerde militære medicinske universitet og Beihang University.
eksperimentelt Design
tre typer arbejdshukommelsesopgaver blev præsenteret for alle deltagere. De var to-back udtale arbejdshukommelsesopgave (se figur 1), to-back rumlig arbejdshukommelsesopgave (se figur 2) og to-back objekt arbejdshukommelsesopgave (se figur 3). Opgaverne stimulerede materialer var 15 store og små bogstaver engelske bogstaver, der ekskluderede dem med lignende bogstaver, såsom L/l, M/m; små sorte firkanter; og 12 geometriske figurer, henholdsvis. Alle materialer blev vist i sort farve på en hvid baggrund med en omtrentlig visuel vinkel på 1,5 liter 1,5 liter (bredde: 2,0 cm, højde: 2,0 cm) subtending. 122 stier blev inkluderet i hver opgave, og i hvert spor blev målstimuleringen præsenteret for 400 ms to stier efter præsentationen af objektiv stimulus med 1.600 ms stimulus start asynkroni tid (SOA), der var præget af en hvid “+.”Deltagerne blev bedt om at klikke på venstre museknap, når målet og objektiv stimulus var den samme (“matchende”), mens du klikker på højre museknap, når de ikke var (“mismatching”). Den matchende eller ikke-betingelse blev præsenteret i en pseudorandom-rækkefølge med et 1:1-forhold.
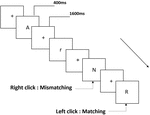
Figur 1. Skematisk diagram over udtalen arbejdshukommelse opgave.
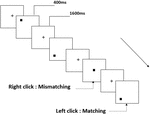
figur 2. Skematisk diagram over den rumlige arbejdshukommelsesopgave.
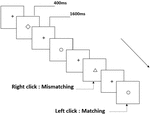
figur 3. Skematisk diagram over objektets arbejdshukommelsesopgave.
eksperimentelle procedurer
før eksperimentet blev deltagerne instrueret af den eksperimentelle opgave. De blev informeret om at øve de tre typer arbejdshukommelsesopgaver, indtil en nøjagtighed på 90% blev opnået. Deltagerne besøgte laboratoriet dagen før eksperimentet og sov i laboratoriet den nat. De to partnerdeltagere udførte eksperimenterne på samme tid. 7: 30 til 8:30 næste morgen med samtidig elektroencefalogram (EEG) optagelse (baseline). Den anden EEG-optagelse (36 h-TSD) blev udført efter en 36-h-periode, hvor deltagerne ikke fik lov til at sove. Under hele eksperimenttiden blev central hæmning og stimulerende stoffer forbudt. Deltagerne blev ledsaget, observeret og mindet af sygeplejepersonale for at holde dem vågen under hele TSD-sessionen.
EEG-optagelser
en kontinuerlig hovedbund EEG blev optaget ved hjælp af elektrodehætter placeret 64 steder ved hjælp af 10-20-systemet med en SynAmps2-forstærker. De bilaterale mastoider (A1 og A2) blev brugt som reference, og panden blev brugt som jord. EEG ‘ er blev registreret ved 1.000 HS, og impedansen af alle kanaler blev opretholdt under 5 k liter. Fire yderligere elektroder blev placeret over og under højre og venstre øjne for at registrere et bipolært lodret og vandret elektrookulogram.
dataanalyse af Adfærdseksperimenter
på grund af tekniske fejl blev to tilfælde slettet, mens andre 14 tilfælde blev inkluderet i den følgende statistiske analyse. Adfærdsdata omfattede den gennemsnitlige reaktionstid, den korrekte hastighed og det korrekte antal pr. Adfærdsmæssige data i baseline og 36 h-TSD-tilstande blev registreret til analyse. Analyserne blev kørt af IBM SPSS (V22.2), hvor metoden gentagne målinger variansanalyse (ANOVA) med drivhus-Geisser var Bonferroni post-hoc-analyse blev lanceret. De statistiske resultater blev præsenteret som middel-og standardafvigelse (SD).
EEG dataanalyse
Scan 4.3-programmet blev brugt til at analysere EEG-dataene, hvor EEG-artefakterne i øjenbevægelsen blev korrigeret ved okulær artefaktreduktionsmetode. Epoker fra -100 til 800 ms af de kontinuerlige EEG-data blev ekstraheret og filtreret af et båndpasfilter fra 0,5 til 30 hs med frekvenshældningen på 24 dB/okt. Forsøgene, hvor spændingen oversteg kr100 kr, blev afvist, og baseline blev korrigeret til en gennemsnitlig amplitude på 100 ms. EEG-komponenterne blev gennemsnitligt og beregnet med kun de korrigerede responser. ERP-komponenterne P2 (100-250 ms), N2 (150-350 ms) og P3 (250-450 ms) i stimulusforsøgene blev identificeret og kvantificeret. De store gennemsnitlige spidsamplituder og latenstider for N2-og P3-komponenterne blev beregnet separat ved F3, FS, F4, C3, C4, P3, PS og P4, og P2-komponenten blev beregnet ved F3, FS, F4, C3, Csog C4 (Casement et al., 2006; Jørgen et al., 2014).
gentagne foranstaltninger ANOVA blev anvendt til alle ERP-resultater. Hovedeffekterne og interaktionerne mellem søvntilstande (baseline og 36 h-TSD), opgaver (udtale arbejdshukommelse, rumlig arbejdshukommelse og objektarbejdshukommelse), regioner (frontal, central og parietal; P2-komponenten blev kun analyseret på de frontale og centrale regioner) og steder (venstre, mellem og højre) blev statistisk analyseret ved hjælp af gentagne målinger ANOVA, som omfattede drivhusgeisser-korrektioner for ikke-sfæricitet og Bonferroni post-hoc-tests.
resultater
adfærdsmæssig ydeevne
resultaterne af adfærdsforsøg er vist i tabel 1. Den gennemsnitlige reaktionstid var længere i 36 h-TSD-tilstanden end ved baseline med en tendens til at stige, men uden signifikante forskelle. ANOVA afslørede, at den korrekte hastighed af opgaven var signifikant forskellig mellem baseline og 36 h-TSD . Tidsenhed viste en signifikant hovedeffekt af tiden under 36 h-TSD .

tabel 1. Ydelsesdata (middelværdi-SD-værdi) på 2-back-opgaven ved baseline og efter 36 h-TSD.
Amplitude
sammenlignet med baseline blev der observeret et signifikant fald i amplituden af P3, og en signifikant stigning blev observeret i amplituden af P2 efter TSD. Selvom N2-amplituden faldt efter 36 h af TSD, nåede forskellen ikke statistisk signifikans (tabel 2).

tabel 2. Grand-gennemsnitlig spidsamplitude af P2 -, N2-og P3-komponenterne i den korrekte responstilstand på tværs af flere elektrodesteder ved baseline og efter 36 h-TSD.
signifikante hovedeffekter af regioner og steder på P2-amplituden blev fundet under TSD-tilstanden. Under TSD optrådte den maksimale amplitude af P2 i frontalområdet (figur 4). Derudover var forskellene i P2-amplituder i forskellige regioner (frontal vs. central) mere signifikante i de bilaterale elektroder (venstre: P = 0,001; højre: P = 0,000) end i de midterste elektroder (figur 4). En signifikant hovedeffekt af regionen og stedet på N2 afslørede, at N2-amplituden var mere negativ i frontal end i den centrale region (P = 0,008, figur 4) og var mindre til højre end på venstre side (P = 0,011,figur 5A2, B2). En hovedvirkning af site på P3 amplitude blev observeret . Amplituden af P3 var mere positiv i midten end på venstre side (P = 0,009, figur 4). En signifikant interaktionseffekt mellem tid og region blev observeret for P3 amplitude . Under TSD var reduktionen i P3-amplitude mere signifikant i de frontale og centrale regioner end i parietalområdet (P = 0,005; P = 0,003) (figur 5C3). Ingen andre hovedeffekter eller interaktionseffekter nåede statistisk signifikans.
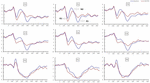
figur 4. ERP-amplitude ved baseline og 36 h-TSD for den korrekte responstilstand for arbejdshukommelsesopgaven. Kanalerne er bestilt fra venstre mod højre og top til bund som følger: F3, FS, F4, C3, C4, P3, PS og P4. Sammenlignet med baseline blev latenserne for N2-P3-komponenterne forlænget, og amplituderne af N2-P3 blev reduceret efter 36 h-TSD.
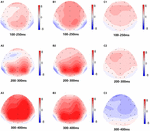
figur 5. Topografisk kort over det korrekte svar i arbejdshukommelsesopgaven under forskellige søvnforhold (A1-C3). (A1) P2, 100-250 ms ved baseline. (A2) N2, 200-300 ms ved baseline. (A3) P3, 300-400 ms ved baseline. (B1) P2, 100-250 ms, ved 36 h-TSD. (B2) N2, 200-300 ms, for 36 h-TSD. (B3) P3, 300-400 ms, til 36 h-TSD. (C1) P2, 100-250 ms, 36 h-TSD med baseline subtraheret. (C2) N2, 200-300 ms, 36 h-TSD med baseline subtraheret. (C3) P3, 300-400 ms, 36 h-TSD med baseline subtraheret.
latens
latenstiden for N2 og P2 blev signifikant forlænget efter TSD. Selvom P3-latensen blev forlænget efter 36 h TSD, nåede forskellen ikke statistisk signifikans (tabel 3).

tabel 3. Grand-gennemsnitlig maksimal latenstid for P2 -, N2-og P3-komponenterne i den korrekte responstilstand på tværs af flere elektrodesteder ved baseline og efter 36 h-TSD.
den signifikante hovedeffekt af region på N2 og P3 afslørede, at latensen af N2-P3-komponenterne var kortere i parietalområdet end i frontalområdet (P = 0,002; P = 0,000) og central region (P = 0,000; P = 0,000) (figur 4). Latensen af P3-bølgen var signifikant længere på venstre side end på højre side (figur 4).
ingen andre hovedeffekter eller interaktionseffekter nåede statistisk signifikans.
N2 -, P2-og P3-amplituder og latenser, der blev fremkaldt på de ni elektrodesteder, er præsenteret i figur 4. Det topografiske kort over det korrekte svar i arbejdshukommelsesopgaven under forskellige søvnforhold (baseline, 36 h-TSD og forskellen mellem de to betingelser) er vist i figur 5.
Diskussion
i denne undersøgelse rapporterede vi påvirkningerne af 36 timers søvnmangel på arbejdshukommelse, der kombinerede adfærdsmæssige data i to søvntilstande (baseline og 36 h-TSD) med samtidige EEG-optagelser. Analysen af resultaterne viste, at ændringerne i adfærdsdataene i overensstemmelse med nedsat arbejdshukommelse efter 36 h TSD: en stigning i den gennemsnitlige reaktionstid for de kognitive opgaver og et fald i nøjagtigheden.
søvnmangel svækkede individets kontrol med Opmærksomme ressourcer. Selvom enkeltpersoner forsøgte at opretholde vågenhed og arbejdsydelse, herunder reaktionstid og korrekt hastighed, under søvnmangel, blev informationsbehandlingskapaciteten i deres arbejdshukommelse stadig påvirket på grund af faldet i hastigheden af behandling af information (Casement et al., 2006; Jørgen et al., 2018). I denne undersøgelse blev N2-og P3-bølgerne relateret til arbejdshukommelse målt for at vise en stigning i latenstid og et fald i amplitude efter søvnmangel sammenlignet med baseline-aflæsningerne. Undersøgelser har vist, at søvnmangel fører til et kontinuerligt fald i opmærksomheden, og fænomenet nedsat P3-amplitude indikerer, at individers top-ned-kontrol af kognition gradvist kollapser. Søvnmangel har en mere negativ indvirkning på kognitive funktioner, især dem, der er afhængige af mentale eller kognitioner., 2019).
P3-komponenten afspejler implementeringen af opmærksomhedsressourcer, og latenstiden for P3 ses bredt som tidsvinduet for stimuluskategorisering og evaluering. Faldet i P3-bølgeamplituden bekræftede også, at beslutningstagningen i det matchende respons, efter at TSD var blevet beskadiget til en vis grad (Gosselin et al., 2005). Undersøgelser har antydet, at søvnmangel kan påvirke informationsbehandlingsstadiet i arbejdshukommelsen. I denne undersøgelse understøttede præstationsindikatorerne også konklusionen om, at responstiden på målstimuleringen blev øget, og at latensen af P3-bølgen blev forlænget (Cote et al., 2008). Det blev spekuleret i, at effekten af søvnmangel på P3-komponenter også kunne finde sted på grund af manglende reaktion på information alter, hvilket er i overensstemmelse med tidligere konklusioner om, at P3-komponenterne er relateret til opdatering af arbejdshukommelsesindhold (Donchin og Fabiani, 1991).
tidligere undersøgelser har betragtet N2-komponenten som et elektrofysiologisk indeks, der afspejler individets evne til at undertrykke responsen (Kreusch et al., 2014). Efter søvnmangel indikerer den langvarige latenstid for NoGo-N2-komponenten, at individets evne til at undertrykke responsen er nedsat (Jin et al., 2015). Den nedsatte amplitude og forlængede latenstid af N2-komponenten relateret til udtale arbejdshukommelse efter søvnmangel afslører, at søvnmangel forringer informationsbehandlingen af udtale arbejdshukommelse (Jhang et al., 2019). N2-komponenten menes generelt at afspejle hjernens selektive opmærksomhed og behandling af følelsesmæssige stimuli eller signaler (Schacht et al., 2008) og er en endogen komponent relateret til en persons mentale tilstand, opmærksomhed og grad af opmærksomhed. I denne undersøgelse fandt vi, at latensen af N2-komponenten var signifikant forlænget, men amplituden viste kun en nedadgående tendens. Ifølge tidligere undersøgelser afspejlede langvarig N2-latenstid en stigning i responstid efter søvnbegrænsning (Jhang et al., 2014). Konstateringen af, at N2-amplitude ikke blev signifikant ændret, kan imidlertid have været på grund af cerebrale kompenserende reaktioner (Drummond og brun, 2001). I tilfælde af begrænsede kognitive ressourcer var der en kompensationsmekanisme til gendannelse af nedsat kognitiv funktion (Jin et al., 2015).
ifølge hovedbundens topografi er ændringerne i N2-P3-komponenterne relateret til søvnmangel mere tydelige i frontalområdet. Frontoparietal kontrol (FPC) spiller en vigtig rolle i kognitiv kontrol. Undersøgelser har vist, at FPC kan omgå kognitiv kontrol ovenfra og ned, hvilket gør det muligt for enkeltpersoner at fokusere på information relateret til målet, mens de undertrykker information, der ikke er relateret til målet., 2011; et al., 2013). FPC er vigtig for opbevaring af information og informationsbehandling i arbejdshukommelsen, og graden af aktivering af FPC efter søvnmangel blev reduceret sammenlignet med en normal søvngruppe (Ma et al., 2014). Selvom EEG-resultaterne ikke afspejlede ændringerne i specifikke hjerneområder i detaljer, afspejlede det intuitivt effekten af TSD på opbevaring og behandling af arbejdshukommelsesinformation.
selvom den nøjagtige kognitive proces, som P2-komponenten ligger til grund for, stadig diskuteres bredt, som en bred definition, afspejler P2-komponenten processen med opmærksomhed og visuel behandling og anses generelt for at være relateret til selektiv opmærksomhed og arbejdshukommelse, hvilket afspejler den tidlige vurdering af den perceptuelle proces (Saito et al., 2001). I denne undersøgelse fandt vi en signifikant stigning i P2-bølgeamplituden efter søvnmangel. Undersøgelser har rapporteret, at P2-bølger, som kan være en del af det tidlige kognitive matchingssystem til meddelelsesbehandling og kan sammenligne sensoriske input med lagret hukommelse (Freunberger et al., 2007) er følsomme over for ændringer i missionens opmærksomhed og krav til arbejdshukommelse (Smith et al., 2002). Funktionel kompensation er en af de unikke funktioner i den menneskelige hjerne og en vigtig faktor for at opretholde kognitiv funktion. Overdreven aktivering af den dorsolaterale præfrontale hjernebark (DLPFC) efter søvnmangel indikerer, at når hjerneressourcerne falder, ser DLPFC ud til at have en kompenserende funktion (Drummond et al., 2004; Choo et al., 2005). Derfor spekulerer vi i, at den signifikante stigning i P2-amplitude observeret i denne undersøgelse kan skyldes funktionel kompensation, hvor individer ser ud til at opretholde normal kognitiv funktion efter søvnmangel. Selvom et stort antal undersøgelser har brugt ERP-teknologi til at undersøge effekten af søvnmangel på kognitive funktioner, er tidlige komponenter som N1 og P2 ikke systematisk undersøgt, og resultaterne er inkonsekvente (Evans and Federmeier, 2007; , 2018; Chang et al., 2019). Der er få undersøgelser undersøge ændringen af P2 komponent under søvnmangel (Mograss et al., 2009). Derfor skal virkningerne af søvnmangel på tidlige komponenter i ERP, såsom P2, stadig undersøges og udforskes yderligere.
i dette eksperiment brugte vi 2-back-modellen til at designe udtale -, rumlige og objektarbejdshukommelsesopgaver og undersøgte forringelsen af arbejdshukommelsen efter 36 h TSD. Sammenlignet med tidligere undersøgelser, der kun fokuserede på virkningerne af søvnmangel på en bestemt type information, såsom udtale arbejdshukommelse eller specifik kognitiv funktion, såsom responshæmning, har vi overvejet indholdet af arbejdshukommelsesmodellen og analyseret grundigt virkningerne af søvnmangel på arbejdshukommelsen.
undersøgelsen har dog nogle begrænsninger. For det første brugte vi kun 2-back-opgaven og kunne ikke sammenligne deltagernes ydeevne i arbejdshukommelsesopgaver med forskellige vanskeligheder. Derfor er der begrænsninger i at forklare og udlede ændringer i arbejdsbyrden. For det andet blev kun mandlige frivillige brugt i undersøgelsen, og konklusionerne skal vurderes, når de udvides til kvindelige frivillige. På grund af det begrænsede antal deltagere fandt vi kun, at amplituden af N2-bølgen havde en nedadgående tendens, og at P3-bølgens latenstid havde en langvarig tendens. Stabile resultater kan opnås efter at have øget antallet af deltagere. For det tredje kan kombination af vores procedure med fMRI til arbejdshukommelse lette yderligere fortolkning af resultaterne. Tidligere undersøgelser har vist, at cirkadiske biorytmer påvirker adfærdsmæssig ydeevne, og der er individuelle forskelle (Montplaisir, 1981; Lavie, 2001). Vi registrerede ikke EEG-dataene på samme tidspunkt i dette eksperiment, så indflydelsen af cirkadiske biorhythmer på testresultaterne kan ikke udelukkes fuldstændigt.
denne undersøgelse viste, at arbejdshukommelsesevnen blev nedsat efter TSD, og at denne skade ikke var forbundet med stimulusindholdet i arbejdshukommelsen. Manglen på søvn reducerede kvaliteten af de oplysninger, der er gemt i hukommelsen, hvilket kan forekomme med den degenerative opmærksomhedsproces (Ratcliff og Van Dongen, 2018). Denne undersøgelse giver elektrofysiologisk bevis for forståelse af mekanismen under nedsat arbejdshukommelse efter søvnmangel. Det er nødvendigt at være opmærksom på de negative virkninger af nedsat arbejdshukommelse forårsaget af søvnmangel og at undersøge effektive indgreb for sådan skade.
Datatilgængelighedserklæring
datasættene genereret til denne undersøgelse er tilgængelige på anmodning til den tilsvarende forfatter.
Etikerklæring
undersøgelserne, der involverede menneskelige deltagere, blev gennemgået og godkendt af det fjerde militære medicinske universitet Beihang University. Patienterne / deltagerne gav deres skriftlige informerede samtykke til at deltage i denne undersøgelse.
Forfatterbidrag
YS designet eksperimenterne. P. P. producerede resultaterne og skrev manuskriptet. CD og LC har analyseret og fortolket data. JT og YS udførte eksperimenterne, kendte dataene og garantisterne for denne undersøgelse. YB, JT og JT bidrog til at deltage i dataindsamling og gennemgang af litteraturen. Alle forfattere, der er anført, har læst og godkendt det endelige manuskript.
finansiering
denne forskning blev støttet af National Science Foundation of Vinter-OL Technology Plan of China under Grant Nos. 2019yff0301600 og HJ20191A020135.
interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
Anderson, C., and Platten, C. R. (2011). Søvnmangel sænker hæmning og forbedrer impulsiviteten til negative stimuli. Opfør dig ordentligt. Brain Res. 217: 466. doi: 10.1016 / j. bbr.2010.09.020
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Baddeley, A. (2000). Den episodiske buffer: en ny komponent i arbejdshukommelsen? Tendenser Cogn. Sci. 4:417. doi: 10.1016 / s1364-6613 (00) 01538-2
CrossRef fuldtekst/Google Scholar
Bartel, P., Offermeier, H., Smith, F. og Becker, P. (2004). Opmærksomhed og arbejdshukommelse hos bosiddende anæstesilæger efter natarbejde: gruppe-og individuelle effekter. Occup. Environ. Middelhavs. 61, 167–170. doi: 10.1136 / oem.2002.006197
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Casement, M. D., Broussard, J. L., Mullington, J. M., and Press, D. Å. (2006). Søvnens Bidrag til forbedringer i arbejdshukommelsesscanningshastighed: en undersøgelse af langvarig søvnbegrænsning. Biol. Psychol. 72:212. doi: 10.1016 / j. biopsycho.2005.11.002
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Chee, M. V. L. og Chuah, L. Y. M. (2008). Funktionel neuroimaging indsigt i, hvordan søvn og søvnmangel påvirker hukommelse og kognition. Curr. Opin. Neurol. 21, 417–423. doi: 10.1097 / toilet.0b013e3283052cf7
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Choo, C. C., Lee, V. V., Sheu, F. S. og Chee, M. V. L. (2005). Dissociation af kortikale regioner moduleret af både arbejdshukommelsesbelastning og søvnmangel og af søvnmangel alene. Neuroimage 25, 579-587. doi: 10.1016 / j.neuroimage.2004.11.029
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Chuah, Y. M. L., Venkatraman, V., Dinges, D. F. og Chee, M. V. (2006). Det neurale grundlag for interindividuel variabilitet i hæmmende effektivitet efter søvnmangel. J. Neurosci. 26, 7156–7162. doi: 10.1523 / jneurosci.0906-06.2006
CrossRef fuldtekst | Google Scholar
Cote, K. A., Milner, C. E., Osip, S. L., Baker, M. L. og Cuthbert, B. P. (2008). Fysiologisk ophidselse og opmærksomhed i løbet af en uge med kontinuerlig søvnbegrænsning. Physiol. Opfør dig ordentligt. 95, 353–364. doi: 10.1016 / j. physbeh.2008.06.016
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Cote, K. A., Mondloch, C. J., Sergeeva, V., Taylor, M. Og Semplonius, T. (2014). Virkningen af total søvnmangel på adfærdsmæssig neural behandling af følelsesmæssigt udtryksfulde ansigter. Eksp. Brain Res. 232, 1429-1442. doi: 10.1007 / s00221-013-3780-1
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Donchin, E., Og Fabiani, M. (1991). Brugen af hændelsesrelaterede hjernepotentialer i studiet af hukommelse: er p300 et mål for begivenheds særpræg? Psychol. Facul. Offentlig. 7, 58–77. doi: 10.1111 / sltb.12055
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Drummond, S. P., brun, G. G., Salamat, J. S. og Gillin, J. C. (2004). Stigende opgavevanskeligheder letter det cerebrale kompenserende respons på total søvnmangel. Sov 27, 445-451. doi: 10.1093 / sleep / 27.3.445
CrossRef Fuld tekst / Google Scholar
Drummond, S. P. A. og brun, G. G. (2001). Virkningerne af total søvnmangel på cerebrale reaktioner på kognitiv præstation. Neuropsykofarmakologi 25, S68-S73. doi: 10.1016 / S0893-133h (01) 00325-6
CrossRef fuldtekst/Google Scholar
Evans, K. M. Og Federmeier, K. D. (2007). Hukommelsen, der er højre og hukommelsen, der er venstre: hændelsesrelaterede potentialer afslører halvkugleformede asymmetrier i kodning og opbevaring af verbal information. Neuropsykologi 45, 1777-1790. doi: 10.1016 / j. neuropsykologi.2006.12.014
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Freunberger, R., Klimesch, H., Doppelmayr, M. og H. Pristller, Y. (2007). Visuel P2 komponent er relateret til Theta faselåsning. Neurosci. LETT. 426, 181–186. doi: 10.1016 / j.neulet.2007.08.062
PubMed Abstract | CrossRef fuldtekst/Google Scholar
g Pristraldine, R., Desgranges, B., Foret, J. og Eustache, F. (2005). Forholdet mellem hukommelsessystemer og søvnstadier. J. Sove Res. 14, 123-140. doi: 10.1111 / j. 1365-2869.2005.00450.PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Gosselin, A., Koninck, J. D. og Campbell, K. B. (2005). Total søvnmangel og nyhedsbehandling: konsekvenser for frontal lobe funktion. Clin. Neurofysiol. 116, 211–222. doi: 10.1016 / j. clinph.2004.07.033
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Harrison, Y. og Horne, J. (1998). Søvntab forringer korte og nye sprogopgaver med et præfrontalt fokus. J. Sove Res. 7, 95-100. doi: 10.1046 / j. 1365-2869.1998. 00104.
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, V. J. Meier, B. (2010). Den samtidige gyldighed af N-back-opgaven som et arbejdshukommelsesmål. Hukommelse 18, 394-412. doi: 10.1080/09658211003702171
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N. og Chamari, K. (2013). Virkningen af delvis søvnmangel på de daglige variationer af kognitiv præstation i uddannede fag. Proc. Soc. Opfør dig ordentligt. Sci. 82, 392–396. doi: 10.1016 / j. sbspro.2013.06.281
CrossRef Fuld tekst / Google Scholar
Jennings, J. R., Munk, T. H. og Molen, M. V. V. D. (2003). Søvnmangel påvirker nogle, men ikke alle processer med tilsynsmæssig opmærksomhed. Psychol. Sci. 14, 473–479. doi: 10.1111/1467-9280.02456
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Jin, J., Ye, E., J., Vang, L. og Lei, Y. (2015). Gendannelsessøvn vender nedsat responshæmning på grund af søvnbegrænsning: bevis fra en visuel begivenhedsrelateret potentialestudie. PLoS One 10:e0142361. doi: 10.1371 / tidsskrift.pone.0142361
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Jones, K., og Harrison, Y. (2001). Frontal lobe funktion, søvn tab og fragmenteret søvn. Sov Med. Åb 5: 475. doi: 10.1053 / smrv.2001.0203
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Koslovsky, M., Og Babkoff, H. (1992). Meta-analyse af forholdet mellem total søvnmangel og ydeevne. Chronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Kreusch, F., E., Vilenne, A. og Hansenne, M. (2014). Alkoholmisbrug og ERP-komponenter i go/no-go-opgaver ved hjælp af alkoholrelaterede stimuli: virkningen af alkoholundgåelse. Int. J. Psychophysiol. 94, 92–99. doi: 10.1016 / j. ijpsycho.2014.08.001
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Kusttor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F. og Huster, R. J. (2019). Søvnmangel påvirker differentielt underkomponenter af kognitiv kontrol. Søvn 42: 016. doi: 10.1093 / sleep / s016
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Lavie, P. (2001). Søvn-vågne som en biologisk rytme. Annu. Pastor Psychol. 52, 277–303. doi: 10.1146 / annurev.psych.52.1.277
CrossRef Fuld tekst / Google Scholar
Lei, Y., Shao, Y., Vang, L., Jai, T. og jou, F. (2015). Storstilet hjernenetværkskobling forudsiger total søvnmangel effekter på kognitiv kapacitet. PLoS en 10: e0133959. doi: 10.1371 / tidsskrift.pone.0133959
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Li, C., Huang, D., Chang, H. og Meng, K. (2016). Korttidshukommelsesunderskud korrelerer med hippocampal-thalamiske funktionelle forbindelsesændringer efter akut søvnbegrænsning. Hjerneafbildning Opfører Sig. 11, 954–963. doi: 10.1007 / s11682-016-9570-1
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Ma, N., Dinges, D. F., Basner, M. og Rao, H. (2014). Hvordan akut total søvntab påvirker den behandlende hjerne: en metaanalyse af neuroimaging-undersøgelser. Sov 38, 233. doi: 10.5665 / søvn.4404
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Miyake, A., og Shah, P. (1999). Modeller af arbejdshukommelse. Mekanismer for aktiv vedligeholdelse og udøvende kontrol. Cambridge: Cambridge University Press.
Google Scholar
Mograss, M. A., Guillem, F., Brassini-Poisson, V. og Godbout, R. (2009). Virkningerne af total søvnmangel på genkendelseshukommelsesprocesser: en undersøgelse af hændelsesrelateret potentiale. Neurobiol. Lære. Mem. 91, 343–352. doi: 10.1016 / j. nlm.2009.01.008
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Montplaisir, J. (1981). Depression og biologisk rytme: terapeutiske virkninger af søvnmangel. L ‘uni l’ uni l ‘uni l’ uni 110, 272-276. doi: 10.1620 / tjem.133.481
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Morris, A. M., So, Y., Lee, K. A., Lash, A. A., og Becker, E.V. T. (1992). Det P300 begivenhedsrelaterede potentiale. virkningerne af søvnmangel. J. Occupat. Middelhavs. 34, 1143–1152.
Google Scholar
Musur, A., Pace-Schott, E. F. og Hobson, J. A. (2002). Den præfrontale bark i søvn. Tendenser Cogn. Sci. 6, 475–481. doi: 10.1016 / S1364-6613 (02) 01992-7
CrossRef fuldtekst | Google Scholar
Ning, M., Dinges, D. F., Mathias, B. Og Hengyi, R. (2014). Hvordan akut total søvntab påvirker den behandlende hjerne: en metaanalyse af neuroimaging-undersøgelser. Sov 38, 233-240. doi: 10.5665 / søvn.4404
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
oven, A. M., Mcmillan, K. M., Laird, A. R. og Bullmore, E. (2005). N-back arbejdshukommelsesparadigme: en metaanalyse af normative funktionelle neuroimaging-studier. Hum. Brain Mapp. 25, 46–59. doi: 10.1002 / hbm.20131
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Panjvani, U., Ray, K., Chatterjee, A., Bhaumik, S. og Kumar, S. (2010). Elektrofysiologiske korrelater af kognition forbedres med lur under søvnmangel. EUR. J. Appl. Physiol. 108, 549–556. doi: 10.1007 / s00421-009-1222-3
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Ratcliff, R. og Van Dongen, H. (2018). Virkningerne af søvnmangel på vare og associativ genkendelseshukommelse. J. Eksp. Psychol. 44, 193–208. doi: 10.1037 / lm0000452
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Raymond, C. A. (1988). Skiftende arbejde, søvncyklusser er på vej til at blive et andet folkesundhedsproblem. JAMA J. Am. Middelhavs. Assoc. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef fuldtekst / Google Scholar
Saito, H., Yamasaki, H., Matsuoka, H., Matsumoto, K. og Sato, M. (2001). Visuel hændelsesrelateret potentiale i mild demens af typen. Psykiatri Clin. Neurosci. 55, 365–371. doi: 10.1046 / j. 1440-1819.2001.00876.
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Schacht, A., Varheid, K. Sommer, M. (2008). Vurderingen af ansigtsskønhed er hurtig, men ikke obligatorisk. Cogn. Påvirke. Opfør dig ordentligt. Neurosci. 8, 132–142. doi: 10.3758 / cabn.8.2.132
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Scott, J. P. R., Mcnaughton, L. R. og Polman, R. C. J. (2006). Effekter af søvnmangel og motion på kognitiv, motorisk ydeevne og humør. Physiol. Opfør dig ordentligt. 87:408. doi: 10.1016 / j. physbeh.2005.11.009
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
lille træ, J., brun, K., Baird, B., Og Schooler, J. V. (2011). Samarbejde mellem standardtilstandsnetværket og det frontale parietale netværk i produktionen af et internt tanketog. Brain Res. 1428, 60-70. doi: 10.1016 / j. brainres.2011.03.072
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Smith, M. E., Mcevoy, L. K. og Gevins, A. (2002). Virkningen af moderat søvntab på neurofysiologiske signaler under arbejdshukommelsesopgavens ydeevne. Sov 25, 784-794. doi: 10.1093/søvn / 25.7.56
CrossRef Fuld tekst / Google Scholar
Thomas, M. (2003). Neuralt grundlag for årvågenhed og kognitive præstationsnedsættelser under søvnighed ii. effekter af 48 og 72 timers søvnmangel på vækkende menneskelig regional hjerneaktivitet. Thalam. Forhold. Syst. 2, 199–229. doi: 10.1016 / S1472-9288 (03) 00020-7
CrossRef fuldtekst | Google Scholar
Van Dongen, H. (2004). De kumulative omkostninger ved yderligere vågenhed: doseresponse virkninger på neuroadfærdsmæssige funktioner og søvnfysiologi fra kronisk søvnmangel og total søvnmangel. Sov 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef Fuld tekst / Google Scholar
Vervij, I. M., Romeijn, N., Smit, D. J. og Giovanni, P. (2014). Søvnmangel fører til et tab af funktionel forbindelse i frontale hjerneområder. BMC Neurosci. 15:88. doi: 10.1186/1471-2202-15-88
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Vi, Vi, Liu, Y., Yao, L. og Ding, M. (2013). Top-ned regulering af standardtilstand aktivitet i rumlig visuel opmærksomhed. J. Neurosci. 33, 6444–6453. doi: 10.1523 / JNEUROSCI.4939-12.2013
CrossRef Fuld tekst | Google Scholar
E., Malayka, M. og Kendra, G. (2018). 24-h søvnmangel forringer tidlig opmærksomhedsmodulation af neural behandling: en begivenhedsrelateret hjernepotentialundersøgelse. Neurosci. LETT. 677, 32–36. doi: 10.1016 / j.neulet.2018.04.022
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Jang, L., Shao, Y., Liu, Li, C., Chen, Y. og Jhou, K. (2019). Nedsat informationsudskiftning af arbejdshukommelse efter søvnmangel: bevis fra en begivenhedsrelateret potentiel undersøgelse. Front. Neurosci. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef Fuld tekst / Google Scholar
Jang, K., Liao, Y., J., J., Y., Y., T., T., Liu, Å, et al. (2014). En visuel ERP-undersøgelse af impulshæmning efter en nap-induceret lur efter søvnmangel. PLoS en 9: e95653. doi: 10.1371 / tidsskrift.pone.0095653
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Leave a Reply