Grenser i Nevrovitenskap
Introduksjon
med samfunnets fremgang og endringer i arbeidsrytmen, lider et økende antall mennesker av søvnmangel. Søvnmangel ikke bare skader den fysiske og mentale helsen til den enkelte, men også alvorlig påvirker arbeid ytelse, forårsaker arbeid feil og selv ulykker. Derfor er forståelse av mekanismen for søvnmangel som påvirker kognitiv funksjon av stor betydning for effektivt å forhindre effekten av søvnmangel.Tidligere studier har vist at søvnmangel kan forårsake en rekke endringer i individets humør, kognitive evne, arbeidsytelse og immunfunksjon (Choo et al., 2005). Mangel på søvn forstyrrer kroppens sirkulasjon og påvirker individers kognitive og følelsesmessige evner (Raymond, 1988). Flere studier har vist at søvnmangel svekker responshemming (Harrison Og Horne, 1998; Muzur et al., 2002; Jennings et al., 2003). For eksempel, etter 36 h søvnmangel, ble individets evne til å undertrykke negative stimuli redusert (Chuah et al ., 2006). Neuroimaging studier har antydet at søvnmangel reduserer individets lave nivå av visuell prosesseringsevne (Anderson og Platten, 2011; Ning et al., 2014). I tillegg svekker søvnmangel hippocampus og kan påvirke minnet ved å ødelegge synaptisk plastisitet (Cote et al., 2014). Thomas (2003) har indikert at mangel på søvn redusert cerebral blodstrøm og metabolisk hastighet i thalamus, prefrontal cortex og parietal cortex (G ④raldine et al., 2005). Jarraya og kolleger fant at delvis søvnmangel påvirket nevropsykologiske funksjoner som verbalt øyeblikkelig minne, oppmerksomhet og våkenhet (Thomas, 2003). Videre har noen studier vist at de kumulative effektene av delvis søvnmangel kan alvorlig svekke kognitiv funksjon og oppførsel (Van Dongen, 2004; Scott et al ., 2006; Jarraya et al., 2013).arbeidsminne Er et system som brukes til å lagre og behandle informasjon, og som er en kognitiv funksjon med begrenset kapasitet (Bartel et al., 2004). Videre kan informasjonen som er lagret i arbeidsminnesystemet endres fra korttidshukommelse til langtidshukommelse gjennom gjenfortelling og andre minnemetoder. Arbeidsminne er overgangen mellom kortsiktige og langsiktige minnesystemer, som er svært sentral i menneskelig meldingsbehandling (Miyake og Shah, 1999). Det gir en midlertidig lagringsplass og ressursene som trengs for å behandle informasjon, for eksempel stemmeforståelse, resonnement og læring. Søvnmangel har vist seg å påvirke arbeidsminnet først.Tidligere studier har brukt n-back arbeidsminneparadigmet hos deltakere som gjennomgikk søvnmangel og fant at mangel på søvn induserer en reduksjon i metabolsk aktivitet i hjernens regionale nettverk, som hovedsakelig utføres informasjonsbehandling og reaksjonshemming (Baddeley, 2000; Zhang et al., 2019). Forringet arbeidsminne etter søvnmangel er relatert til aktiveringen av standardnettverket i oppgaver (Chee og Chuah, 2008), som kan være relatert til thalamus viktige rolle i kortikal våkenhet. For eksempel økte søvnmangel forbindelsen mellom hippocampus, thalamus og standardnettverket, som ofte ble ledsaget av høyere subjektiv døsighet og dårligere ytelse av arbeidsminne (Lei et al., 2015; Li et al., 2016). Studier på søvnmangel identi?ed at økt latens og redusert amplitude Av p3 komponent var assosiert med langvarig nøkternhet (Morris et al., 1992; Jones og Harrison, 2001; Panjwani et al., 2010). Nedgangen I p3-bølgen kan gjenspeile en nedgang i deltakernes oppmerksomhet og en reduksjon i dømmekraften av målstimuli (Koslowsky og Babkoff, 1992).
imidlertid har få studier gitt elektrofysiologiske bevis for nedsatt arbeidsminne etter søvnmangel. N-back-oppgaven regnes som en vanlig metode for å vurdere arbeidsminne (Owen et al., 2005; Jaeggi et al., 2010). Zhang et al. designet en to-back uttale arbeidsminneoppgave for å utforske den reduserte meldingsalternativet for arbeidsminne under søvnmangel, men få studier har brukt forskjellige typer arbeidsminneoppgaver i et enkelt eksperiment. I denne studien utviklet vi ulike typer arbeidsminneoppgaver (uttale arbeidsminne, romlig arbeidsminne og objektarbeidsminne) for å utforske nedsatt kognitiv funksjon AV TSD og registrert deltaker EEG-data ved 2 tidspunkter (baseline og 36 h-TSD). Alle oppgavene vedtatt en 2-back paradigme. Denne studien evaluerte endringene I n2-P3-bølgen relatert til arbeidsminne under TSD og analyserte de tidsmessige egenskapene til effekten av søvnmangel på arbeidsminne. Våre funn gir eksperimentelle bevis for effekten av søvnmangel på kognitiv funksjon.
Materialer og Metoder
Deltakere
Seksten unge, friske, høyrehendte mannlige studenter deltok i denne studien. Vi rekrutterte deltakere ved å annonsere på campus. Deltakerne hadde alle gode søvnvaner(PSQI < 5). Alle deltakerne var mellom 21 og 28 år med en gjennomsnittsalder på 23 år, og ingen av deltakerne hadde noen psykisk eller fysisk sykdom. Alle deltakerne hadde normalt syn eller korrigert syn over 1,0 og intelligenspoeng >110 på Raven-Testen. Før forsøket forklarte eksperimentøren prosedyren og peker på oppmerksomheten til deltakerne for å sikre at de var kjent med metoden og prosedyren. I de 2 ukene før forsøket sov deltakerne regelmessig for 7-9 h per dag uten å røyke, drikke kaffe, drikke alkohol eller konsumere medisiner i 2 dager før forsøket. Før forsøket ga alle deltakerne skriftlig informert samtykke. Forsøksordningen ble godkjent Av Etikkutvalget For Det Fjerde Militære Medisinske Universitetet og Beihang University.
Eksperimentell Design
Tre typer arbeidsminneoppgaver ble presentert for alle deltakere. De var to-back uttale arbeidsminneoppgave (Se Figur 1), to-back romlig arbeidsminneoppgave (Se Figur 2) og to-back objekt arbeidsminneoppgave (Se Figur 3). De stimulere materialer av oppgavene var 15 små bokstaver engelske bokstaver som unntatt de med lignende bokstaver, for Eksempel L/l, M/m; små svarte firkanter; og 12 geometriske figurer, henholdsvis. Alle materialene ble vist i svart farge på hvit bakgrunn, med en omtrentlig visuell vinkel på 1.5 ° × 1.5° (bredde: 2.0 cm, høyde: 2.0 cm) subtending. 122 stier besto av hver oppgave, og i hver sti ble målstimulansen presentert for 400 ms to stier etter presentasjonen av objektiv stimulus, med 1600 ms stimulusutbrudd asynkron tid (SOA) som var preget av en hvit «+.»Deltakerne ble bedt om å klikke på venstre museknapp når målet og målet stimulus var den samme («matching»), mens klikk på høyre museknapp når de ikke var («mismatching»). Samsvarende eller ikke-tilstanden ble presentert i en pseudorandom rekkefølge med et 1: 1-forhold.
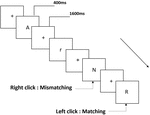
Figur 1. Skjematisk diagram av uttalen arbeidsminne oppgave.
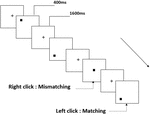
Figur 2. Skjematisk diagram av romlig arbeidsminneoppgave.
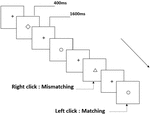
Figur 3. Skjematisk diagram av objektet arbeidsminne oppgave.
Eksperimentelle Prosedyrer
før forsøket ble deltakerne instruert om eksperimentell oppgave. De ble informert om å praktisere de tre typer arbeidsminneoppgaver til en nøyaktighetsgrad på 90% ble oppnådd. Deltakerne besøkte laboratoriet dagen før forsøket og sov i laboratoriet den kvelden. De to partnerdeltakerne utførte forsøkene samtidig. Tre typer arbeidsminneoppgaver ble utført klokken 7: 30 til 8: 30 neste morgen med samtidig elektroensfalogram (EEG) opptak (baseline). Det andre eeg-opptaket (36 h-TSD) ble utført etter en 36-h periode hvor deltakerne ikke fikk lov til å sove. Under hele eksperimenttiden ble sentral hemming og stimulerende stoffer forbudt. Deltakerne ble ledsaget, observert og minnet av pleiepersonalet for å holde dem våken gjennom TSD-økten.
Eeg Opptak
en kontinuerlig hodebunn EEG ble tatt opp ved hjelp av elektrode caps plassert i 64 steder ved hjelp av 10-20 system Med En SynAmps2 forsterker. De bilaterale mastoidene (A1 og A2) ble brukt som referanse, og pannen ble brukt som bakken. Eeg ble registrert ved 1000 Hz, og impedansen til alle kanaler ble opprettholdt under 5 kΩ. Fire ekstra elektroder ble plassert over og under høyre og venstre øyne for å registrere et bipolar vertikalt og horisontalt elektrookulogram.
Dataanalyse Av Atferdsforsøk
på grunn av tekniske feil ble to tilfeller slettet, mens andre 14 tilfeller ble inkludert i følgende statistiske analyse. Atferdsdata inkluderte gjennomsnittlig reaksjonstid, riktig hastighet og riktig antall per tidsenhet. Adferdsdata i baseline og 36 h-TSD-tilstander ble registrert for analyse. Analysene ble utført AV IBM SPSS (V22).2), hvor den gjentatte tiltak analyse av varians (ANOVA) metode med Drivhus-Geisser Var Bonferroni post-hoc analyse ble lansert. De statistiske resultatene ble presentert som gjennomsnitt og standardavvik (SD).
Eeg Data Analysis
Scan 4.3-programmet ble brukt til å analysere EEG-dataene, hvor eeg-artefakter av øyebevegelsen ble korrigert ved okulær artefaktreduksjonsmetode. Epoker fra -100 til 800 ms av de kontinuerlige eeg-dataene ble ekstrahert og filtrert av et båndpassfilter fra 0,5 til 30 Hz med frekvenshellingen på 24 dB/okt. Studiene der spenningen oversteg ± 100 µ ble avvist og baseline ble korrigert til en gjennomsnittlig amplitude på 100 ms. EEG-komponentene ble gjennomsnittlig og beregnet med bare de korrigerte svarene. ERP-komponentene P2 (100-250 ms), N2 (150-350 ms) og P3 (250-450 ms) i stimulusforsøkene ble identifisert og kvantifisert. De store gjennomsnittlige toppamplitudene og latensene Til n2-og p3-komponentene ble beregnet separat Ved F3, Fz, F4, C3, Cz, C4, P3, Pz og P4, og p2-komponenten ble beregnet Ved F3, Fz, F4, C3, Cz og C4 (Casement et al., 2006; Verweij et al., 2014).
Gjentatte tiltak ANOVA ble ansatt for ALLE ERP-resultater. De viktigste effektene og samspillet mellom søvntilstander (baseline og 36 h-TSD), oppgaver (uttale arbeidsminne, romlig arbeidsminne og objektarbeidsminne), regioner (frontal, sentral og parietal; p2-komponenten ble analysert bare på frontal og sentrale regioner) og steder (venstre, midt og høyre) ble statistisk analysert ved bruk av gjentatte tiltak ANOVA, som inkluderte Drivhus-Geisser korreksjoner for ikke-sfærisk og Bonferroni post-hoc tester.
Resultater
Atferdsmessig Ytelse
resultatene av atferdsforsøkene er vist I Tabell 1. Gjennomsnittlig reaksjonstid var lengre i 36 h-TSD-tilstanden enn ved baseline med en trend å øke, men uten signifikante forskjeller. ANOVA viste at den riktige frekvensen av oppgaven var signifikant forskjellig mellom grunnlinjen og 36 h-TSD . Det riktige tallet per tidsenhet viste en signifikant hovedeffekt av tid under 36 h-TSD .

Tabell 1. Ytelsesdata (gjennomsnittlig ± SD) på 2-bakoppgaven ved baseline og etter 36 h-TSD.
Amplitude
Sammenlignet med baseline ble det observert en signifikant reduksjon I amplitude Av P3, og en signifikant økning ble observert I amplitude Av P2 etter TSD. Selv om n2 amplitude redusert etter 36 h AV TSD, forskjellen ikke nå statistisk signifikans (Tabell 2).

Tabell 2. Grand-gjennomsnittlig toppamplitude Av p2 -, N2-og P3-komponentene i riktig responstilstand på tvers av flere elektrodesteder ved baseline og etter 36 h-TSD.
Signifikante hovedeffekter av regioner og steder på p2-amplituden ble funnet under TSD-tilstanden. UNDER TSD oppstod maksimal amplitude Av P2 i frontalområdet(Figur 4). I Tillegg var forskjellene I p2-amplituder i forskjellige regioner (frontal vs sentral) mer signifikante i de bilaterale elektrodene (venstre: P = 0,001; høyre: P = 0,000) enn i de midterste elektrodene (Figur 4). En signifikant hovedvirkning av regionen og stedet På N2 viste at n2 amplitude var mer negativ i frontal enn i sentralområdet (P = 0,008, Figur 4) og var mindre til høyre enn på venstre side (P = 0,011, Figurer 5A2,B2). En hovedeffekt av stedet på p3 amplitude ble observert . Amplituden Til P3 var mer positiv i midten enn på venstre side (P = 0,009, Figur 4). En signifikant interaksjonseffekt mellom tid og region ble observert For p3-amplituden . UNDER TSD var reduksjonen I p3 amplitude mer signifikant i frontale og sentrale områder enn i parietalområdet (P = 0,005; P = 0,003) (Figur 5c3). Ingen andre hovedeffekter eller interaksjonseffekter nådde statistisk signifikans.
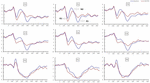
Figur 4. Erp amplitude ved baseline og 36 h-TSD for riktig responsbetingelse for arbeidsminneoppgaven. Kanalene er bestilt fra venstre til høyre og topp til bunn som følger: F3, Fz, F4, C3, Cz, C4, P3, Pz Og P4. Sammenlignet med baseline ble latensene av n2-P3-komponentene forlenget, og amplitudene Til N2-P3 ble redusert etter 36 h-TSD.
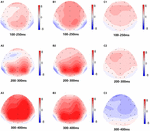
Figur 5. Topografisk kart over riktig respons i arbeidsminneoppgaven i forskjellige søvnforhold(A1-C3). (A1) P2, 100-250 ms, ved baseline. (A2) N2, 200-300 ms, ved baseline. (A3) P3, 300-400 ms, ved baseline. (B1) P2, 100-250 ms, ved 36 h-TSD. (B2) N2, 200-300 ms, for 36 h-TSD. (B3) P3, 300-400 ms, for 36 h-TSD. (C1) P2, 100-250 ms, 36 h-TSD med baseline trukket fra. (C2) N2, 200-300 ms, 36 h-TSD med baseline trukket fra. (C3) P3, 300-400 ms, 36 h-TSD med baseline trukket fra.
Latens
latensene Til N2 Og P2 ble signifikant forlenget etter TSD. Selv Om p3-latensen ble forlenget etter 36 timer MED TSD, nådde ikke forskjellen statistisk signifikans(Tabell 3).

Tabell 3. Gjennomsnittlig maksimal ventetid For p2 -, N2-og P3-komponentene i riktig responstilstand på tvers av flere elektrodesteder ved baseline og etter 36 h-TSD.den signifikante hovedvirkningen av regionen På N2 Og P3 viste at latensen Til n2-P3-komponentene var kortere i parietalområdet enn i frontalområdet (P = 0,002; P = 0,000) Og sentralregionen (P = 0,000; P = 0,000) (Figur 4). Latensen Til p3-bølgen var betydelig lengre på venstre side enn på høyre side(Figur 4).
ingen andre hovedeffekter eller interaksjonseffekter nådde statistisk signifikans.
n2, P2 og P3 amplituder og ventetider som ble fremkalt på de ni elektrodestedene er presentert i Figur 4. Det topografiske kartet over riktig respons i arbeidsminneoppgaven i forskjellige søvnforhold (baseline, 36 h-TSD og forskjellen mellom De to forholdene) er presentert i Figur 5.
Diskusjon
i denne studien rapporterte vi påvirkningen av 36 h søvnmangel på arbeidsminne, og kombinerte atferdsdata i to søvntilstander (baseline og 36 h-TSD) med samtidige eeg-opptak. Analysen av resultatene indikerte at endringene i atferdsdata i samsvar med nedsatt arbeidsminne etter 36 h TSD: en økning i gjennomsnittlig reaksjonstid for kognitive oppgaver og en reduksjon i nøyaktighet.
Søvnmangel svekket individets kontroll over oppmerksomhetsressurser. Selv om enkeltpersoner forsøkte å opprettholde våkenhet og arbeidsytelse, inkludert reaksjonstid og korrekt hastighet, under søvnmangel, ble informasjonsbehandlingskapasiteten til arbeidsminnet fortsatt påvirket på grunn av nedgangen i behandlingshastigheten (Casement et al., 2006; Wiggins et al., 2018). I denne studien ble n2-og P3-bølgene relatert til arbeidsminne målt for å vise en økning i latens og en reduksjon i amplitude etter søvnmangel sammenlignet med baseline-avlesningene. Studier har vist at søvnmangel fører til en kontinuerlig nedgang i oppmerksomheten, og fenomenet redusert p3 amplitude indikerer at individers topp-down kontroll av kognisjon gradvis kollapser. Søvnmangel har en mer negativ effekt på kognitive funksjoner, spesielt de som er avhengige av mental eller kognisjon (Kusztor et al., 2019).
p3-komponenten gjenspeiler distribusjon av oppmerksomhetsressurser, og ventetiden Til P3 er allment sett som tidsvinduet for stimuluskategorisering og evaluering. Nedgangen I p3-bølgeamplituden bekreftet også at beslutningsprosessen i samsvarende respons etter TSD hadde blitt skadet til en viss grad (Gosselin et al., 2005). Studier har antydet at søvnmangel kan påvirke informasjonsbehandlingsstadiet i arbeidsminnet. I denne studien støttet resultatindikatorene også konklusjonen om at responstiden til målstimulansen ble økt, og at latensen Til p3-bølgen var forlenget (Cote et al., 2008). Det ble spekulert i at effekten av søvnmangel på p3-komponenter også kan skje på grunn av manglende respons på informasjonsalter, noe som stemmer overens med tidligere konklusjoner om At P3-komponentene er relatert til oppdatering av arbeidsminneinnhold (Donchin Og Fabiani, 1991).
Tidligere studier har vurdert n2-komponenten som en elektrofysiologisk indeks som reflekterer individets evne til å undertrykke responsen (Kreusch et al., 2014). Etter søvnmangel indikerer langvarig latens Av nogo-N2-komponenten at individets evne til å undertrykke responsen er svekket (Jin et al., 2015). Den reduserte amplitude Og langvarig latens Av n2-komponenten relatert til uttale arbeidsminne etter søvnmangel avslører at søvnmangel forringer informasjonsbehandling av uttale arbeidsminne (Zhang et al., 2019). N2-komponenten er generelt antatt å reflektere hjernens selektive oppmerksomhet og behandling av emosjonelle stimuli eller signaler (Schacht et al., 2008) og er en endogen komponent relatert til individets mentale tilstand, oppmerksomhet og grad av oppmerksomhet. I denne studien fant vi at latensen Til n2-komponenten var betydelig forlenget, men amplituden viste bare en nedadgående trend. Ifølge tidligere studier reflekterte langvarig n2-latens en økning i responstid etter søvnbegrensning (Zhang et al., 2014). Funnet om At n2 amplitude ikke var signifikant endret, kan imidlertid ha vært på grunn av cerebral kompenserende responser (Drummond and Brown, 2001). I tilfelle av begrensede kognitive ressurser var det en kompensasjonsmekanisme for å gjenopprette nedsatt kognitiv funksjon (Jin et al., 2015).
ifølge hodebunnstopografien er endringene I n2-P3-komponentene relatert til søvnmangel mer åpenbare i frontområdet. Frontoparietal kontroll (fpc) spiller en viktig rolle i kognitiv kontroll. Studier har vist AT FPC kan omgå top-down kognitiv kontroll, slik at enkeltpersoner kan fokusere på informasjon relatert til målet mens undertrykke informasjon som ikke er relatert til målet (Smallwood et al., 2011; Wen et al., 2013). FPC er viktig for oppbevaring av informasjon og informasjonsbehandling i arbeidsminnet, og graden AV aktivering AV FPC etter søvnmangel ble redusert sammenlignet med en normal søvngruppe (Ma et al ., 2014). SELV OM eeg-resultatene ikke reflekterte endringene i bestemte hjernegrupper i detalj, reflekterte det intuitivt EFFEKTEN AV TSD på oppbevaring og behandling av arbeidsminneinformasjon.
Selv om den nøyaktige kognitive prosessen Som p2-komponenten ligger til grunn for, fortsatt diskuteres mye, som en bred definisjon, Reflekterer p2-komponenten prosessen med oppmerksomhet og visuell prosessering og anses generelt å være relatert til selektiv oppmerksomhet og arbeidsminne, noe som reflekterer den tidlige dommen av den perceptuelle prosessen (Saito et al., 2001). I denne studien fant vi en signifikant økning I p2-bølgeamplituden etter søvnmangel. Studier har rapportert At p2 bølger som kan være en del av den tidlige kognitive matching system for meldingsbehandling og kan sammenligne sensoriske innganger til lagret minne (Freunberger et al., 2007) er følsomme for endringer i oppdrag oppmerksomhet og arbeidsminne krav (Smith et al., 2002). Funksjonell kompensasjon er en av de unike funksjonene i den menneskelige hjerne og en viktig faktor for å opprettholde kognitiv funksjon. Overdreven aktivering av dorsolateral prefrontal cortex (DLPFC) etter søvnmangel indikerer at ETTER hvert som hjerneressursene reduseres, SER DLPFC ut til å ha en kompenserende funksjon (Drummond et al., 2004; Choo et al., 2005). Derfor spekulerer vi på at den signifikante økningen I p2 amplitude observert i denne studien kan skyldes funksjonell kompensasjon der individer ser ut til å opprettholde normal kognitiv funksjon etter søvnmangel. Selv om et stort antall studier har brukt ERP-teknologi for å undersøke effekten av søvnmangel på kognitive funksjoner, har tidlige komponenter Som N1 Og P2 ikke blitt systematisk studert, og resultatene er inkonsekvente (Evans Og Federmeier, 2007; Wiggins et al., 2018; Zhang et al., 2019). Det er få forskere som undersøker endringen Av P2-komponent under søvnmangel (Mograss et al., 2009). Derfor må effekten av søvnmangel på tidlige komponenter I ERP, som P2, fortsatt studeres og utforskes.I dette eksperimentet brukte vi 2-bakmodellen til å designe uttale, romlig og objektarbeidsminneoppgaver og undersøkte forringelsen av arbeidsminne etter 36 h AV TSD. Sammenlignet med tidligere studier som bare fokuserte på effekten av søvnmangel på en bestemt type informasjon, for eksempel uttale arbeidsminne eller spesifikk kognitiv funksjon, for eksempel responshemming, har vi vurdert innholdet i arbeidsminnemodellen og analysert effekten av søvnmangel på arbeidsminne.
studien har imidlertid noen begrensninger. Først brukte vi bare 2-back-oppgaven og klarte ikke å sammenligne deltakernes ytelse i arbeidsminneoppgaver med ulike vanskeligheter. Derfor er det begrensninger i å forklare og utlede endringer i arbeidsbelastning. For det andre ble bare mannlige frivillige brukt i studien, og konklusjonene må vurderes når de utvides til kvinnelige frivillige. På grunn av det begrensede antall deltakere fant vi bare at amplituden Til n2-bølgen hadde en nedadgående trend, Og At p3-bølgeforsinkelsen hadde en langvarig trend. Stabile resultater kan oppnås etter å ha økt antall deltakere. For det tredje kan kombinere vår prosedyre med fMRI for arbeidsminne lette videre tolkning av resultatene. Tidligere studier har vist at sirkadiske biorhythmer påvirker atferdsytelse ,og det er individuelle forskjeller (Montplaisir, 1981; Lavie, 2001). VI registrerte IKKE EEG-dataene samtidig i denne eksperimentelle, så påvirkning av sirkadiske biorhythmer på testresultatene kan ikke helt utelukkes.denne undersøkelsen viste at arbeidsminneevnen var svekket etter TSD, og at denne skaden ikke var forbundet med stimulansinnholdet i arbeidsminnet. Mangel på søvn reduserte kvaliteten på informasjonen som er lagret i minnet, noe som kan oppstå med degenerativ oppmerksomhetsprosess (Ratcliff Og Van Dongen, 2018). Denne studien gir elektrofysiologi bevis for å forstå mekanismen under nedsatt arbeidsminne etter søvnmangel. Det er nødvendig å være oppmerksom på de negative effektene av arbeidsminneforringelse forårsaket av søvnmangel og å utforske effektive tiltak for slike skader.
Datatilgjengelighetserklæring
datasettene som genereres for denne studien, er tilgjengelige på forespørsel til den tilsvarende forfatteren.
Etisk Erklæring
studiene som involverte menneskelige deltakere ble gjennomgått og godkjent av Det Fjerde Militære Medisinske Universitetet Beihang University. Pasientene / deltakerne ga skriftlig informert samtykke til å delta i denne studien.
Forfatterbidrag
ys designet forsøkene. ZP produserte resultatene og skrev manuskriptet. CD og LZ analyserte og tolket dataene. JT og YS utførte forsøkene, kjente dataene og garantistene for denne studien. YB, LZ og JT bidro til å delta i datainnsamling og gjennomgang av litteraturen. Alle forfatterne har lest og godkjent det endelige manuskriptet.
Finansiering
denne forskningen ble støttet Av National Science Foundation Of Winter Olympics Technology Plan Of China Under Grant Nos. 2019yff0301600 OG HJ20191A020135.
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Anderson, C., Og Platten, C. R. (2011). Søvnmangel reduserer hemming og øker impulsiviteten til negative stimuli. Oppfør deg. Hjernen Res. 217: 466. doi: 10.1016 / j.bbr.2010.09.020
PubMed Abstract | CrossRef Full Text | Google Scholar
Baddeley, a. (2000). Episodisk buffer: en ny komponent i arbeidsminnet? Trender Cogn. Sci. 4:417. doi: 10.1016 / s1364-6613(00)01538-2
CrossRef Full Text | Google Scholar
Bartel, P., Offermeier, W., Smith, F. og Becker, P. (2004). Oppmerksomhet og arbeidsminne hos fastboende anestesileger etter nattevakt: gruppe-og individuelle effekter. Occup. Miljø. Med. 61, 167–170. doi: 10.1136 / oem.2002.006197
PubMed Abstract / CrossRef Full Text / Google Scholar
Casement, Md, Broussard, Jl, Mullington, Jm, Og Press, Dz (2006). Søvnens bidrag til forbedringer i arbeidsminnets skannehastighet: en studie av langvarig søvnbegrensning. Biol. Psychol. 72:212. doi: 10.1016 / j.biopsycho.2005.11.002
PubMed Abstract / CrossRef Full Text / Google Scholar
Chee, M. W. L., Og Chuah, L. Y. M. (2008). Funksjonell neuroimaging innsikt i hvordan søvn og søvnmangel påvirker minne og kognisjon. Curr. Opin. Neurol. 21, 417–423. doi: 10.1097 / WCO.0b013e3283052cf7
PubMed Abstract | CrossRef Full Text / Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, Fs, Og Chee, M. W. L. (2005). Dissosiasjon av kortikale regioner modulert av både arbeidsminnebelastning og søvnmangel og ved søvnmangel alene. Neuroimage 25, 579-587. doi: 10.1016 / j. neuroimage.2004.11.029
PubMed Abstract | CrossRef Full Text | Google Scholar
Chuah, Yml, Venkatraman, V., Dinges, Df, Og Chee, Mwl (2006). Det nevrale grunnlaget for interindividuell variabilitet i hemmende effektivitet etter søvnmangel. J. Neurosci. 26, 7156–7162. doi: 10.1523 / jneurosci.0906-06. 2006
CrossRef Full Text | Google Scholar
Cote, Ka, Milner, Ce, Osip, Sl, Baker, Ml, Og Cuthbert, Bp (2008). Fysiologisk opphisselse og oppmerksomhet i løpet av en uke med kontinuerlig søvnbegrensning. Physiol. Oppfør deg. 95, 353–364. doi: 10.1016 / j. physbeh.2008.06.016
PubMed Abstract | CrossRef Full Text/Google Scholar
Cote, K. A., Mondloch, C. J., Sergeeva, V., Taylor, M., Og Semplonius, T. (2014). Effekt av total søvnmangel på atferdsmessig nevralbehandling av følelsesmessig uttrykksfulle ansikter. Exp. Brain Res. 232, 1429-1442. doi: 10.1007 / s00221-013-3780-1
PubMed Abstract | CrossRef Full Text/Google Scholar
Donchin, E., Og Fabiani, M. (1991). Bruken av hendelsesrelaterte hjernepotensialer i studiet av minne: er p300 et mål for hendelses egenart? Psychol. Facul. Offentlig. 7, 58–77. doi: 10.1111 / sltb.12055
PubMed Abstract | CrossRef Full Text/Google Scholar
Drummond, S. P., Brown, G. G., Salamat, J. S., og Gillin, J. C. (2004). Økende oppgave vanskeligheter letter cerebral kompenserende respons på total søvnmangel. Sov 27, 445-451. doi: 10.1093 / søvn / 27.3.445
CrossRef Full Text / Google Scholar
Drummond, Spaog Brown, Gg (2001). Effektene av total søvnmangel på cerebrale responser på kognitiv ytelse. Neuropsykofarmakologi 25, S68-S73. doi: 10.1016 / S0893-133x (01) 00325-6
CrossRef Full Text/Google Scholar
Evans, K. M. Og Federmeier, K. D. (2007). Minnet som er høyre og minnet som er venstre: hendelsesrelaterte potensialer avslører hemisfæriske asymmetrier i koding og oppbevaring av verbal informasjon. Neuropsykologi 45, 1777-1790. doi: 10.1016 / j. nevropsykologi.2006.12.014
PubMed Abstract | CrossRef Full Text/Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M., Og Hö, Y. (2007). Visuell p2-komponent er relatert til theta-faselåsing. Neurosci. Lett. 426, 181–186. doi: 10.1016 / j.neulet.2007.08.062
PubMed Abstract | CrossRef Full Text/Google Scholar
Gé, R., Desgranges, B., Foret, J., Og Eustache, F. (2005). Forholdet mellom hukommelsessystemer og søvnstadier. J. Sleep Res. 14, 123-140. doi: 10.1111/j.1365-2869. 2005. 00450.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Gosselin, A., Koninck, J. D., Og Campbell, K. B. (2005). Total søvnmangel og novelty processing: implikasjoner for frontal lobe fungerer. Clin. Nevrofysiol. 116, 211–222. doi: 10.1016 / j.clinph.2004.07.033
PubMed Abstract | CrossRef Full Text / Google Scholar
Harrison, Y., Og Horne, J. (1998). Søvnmangel svekker korte og nye språkoppgaver som har et prefrontalt fokus. J. Sleep Res. 7, 95-100. doi: 10.1046/j.1365-2869. 1998. 00104.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J. Og Meier, B. (2010). Den samtidige gyldigheten av n-back-oppgaven som et arbeidsminnemål. Minne 18, 394-412. doi: 10.1080/09658211003702171
PubMed Abstract | CrossRef Full Text/Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., Og Chamari, K. (2013). Virkningen av delvis søvnmangel på de daglige variasjonene av kognitiv ytelse hos trente fag. Proc. Soc. Oppfør deg. Sci. 82, 392–396. doi: 10.1016 / j. sbspro.2013.06.281
CrossRef Full Text/Google Scholar
Jennings, J. R., Monk, T. H., og Molen, M. W. V. D. (2003). Søvnmangel påvirker noen, men ikke alle, prosesser med tilsynsoppmerksomhet. Psychol. Sci. 14, 473–479. doi: 10.1111/1467-9280.02456
PubMed Abstract | CrossRef Full Text | Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L. og Lei, Y. (2015). Recovery sleep reverserer nedsatt responshemming på grunn av søvnbegrensning: bevis fra en visuell hendelsesrelatert potensialstudie. PLoS One 10:e0142361. doi: 10.1371 / tidsskrift.pone.0142361
PubMed Abstract | CrossRef Full Text/Google Scholar
Jones, K., Og Harrison, Y. (2001). Frontal lobe funksjon, søvn tap og fragmentert søvn. Sov Med. Åp 5: 475. doi: 10.1053 / smrv.2001.0203
PubMed Abstract | CrossRef Full Text/Google Scholar
Koslowsky, M., Og Babkoff, H. (1992). Meta-analyse av forholdet mellom total søvnmangel og ytelse. Chronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
PubMed Abstract | CrossRef Full Text/Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A., Og Hansenne, M. (2014). Alkoholmisbruk og erp-komponenter i go / no-go-oppgaver ved bruk av alkoholrelaterte stimuli: påvirkning av alkoholunngåelse. Int. J. Psykofysiol. 94, 92–99. doi: 10.1016 / j. ijpsycho.2014.08.001
PubMed Abstract | CrossRef Full Text/Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F., Og Huser, R. J. (2019). Søvnmangel påvirker differensielt underkomponenter av kognitiv kontroll. Sove 42: zsz016. doi: 10.1093 / søvn/zsz016
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Lavie, P. (2001). Søvn-våkne som en biologisk rytme. Annu. Pastor Psychol. 52, 277–303. doi: 10.1146 / annurev.psych.52.1.277
CrossRef Full Text | Google Scholar
Lei, Y., Shao, Y., Wang, L., Zhai, T., Og Zou, F. (2015). Storskala hjernenettverkskobling spår total søvnmangel effekter på kognitiv kapasitet. PLoS En 10: e0133959. doi: 10.1371 / tidsskrift.pone.0133959
PubMed Abstract | CrossRef Full Text/Google Scholar
Li, C., Huang, D., Qi, J., Chang, H. og Meng, Q. (2016). Kortsiktige minne underskudd korrelerer med hippocampal-thalamic funksjonelle tilkobling endringer etter akutt søvn begrensning. Brain Imaging Oppfører Seg. 11, 954–963. doi: 10.1007 / s11682-016-9570-1
PubMed Abstract | CrossRef Full Text/Google Scholar
Ma, N., Dinges, Df, Basner, M., Og Rao, H. (2014). Hvordan akutt totalt søvnfall påvirker den tilstedeværende hjernen: en meta-analyse av neuroimaging studier. Sov 38, 233. doi: 10.5665 / søvn.4404
PubMed Abstract | CrossRef Full Text/Google Scholar
Miyake, A., Og Shah, P. (1999). Modeller Av Arbeidsminne. Mekanismer For Aktivt Vedlikehold og Utøvende Kontroll. Cambridge: Cambridge University Press.S.
Google Scholar
Mograss, Ma, Guillem, F., Brazzini-Poisson, V., Og Godbout, R. (2009). Effektene av total søvnmangel på anerkjennelsesminneprosesser: en studie av hendelsesrelatert potensial. Neurobiol. Lære. Mem. 91, 343–352. doi: 10.1016 / j. nlm.2009.01.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Montplaisir, J. (1981). Depresjon og biologisk rytme: terapeutiske effekter av søvnmangel. L ‘ unió Mé i Canada 110, 272-276. doi: 10.1620 / hjem.133.481
PubMed Abstract | CrossRef Full Text/Google Scholar
Morris, A. M., So, Y., Lee, K. A., Lash, A. A., Og Becker, C. E. (1992). Det p300 hendelsesrelaterte potensialet. effektene av søvnmangel. J. Occupat. Med. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F., Og Hobson, J. A. (2002). Den prefrontale cortex i søvn. Trender Cogn. Sci. 6, 475–481. doi: 10.1016 / S1364-6613(02)01992-7
CrossRef Full Text | Google Scholar
Ning, M., Dinges, Df, Mathias, B. og Hengyi, R. (2014). Hvordan akutt totalt søvnfall påvirker den tilstedeværende hjernen: en meta-analyse av neuroimaging studier. Sov 38, 233-240. doi: 10.5665 / søvn.4404
PubMed Abstract | CrossRef Full Text/Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, Ar, Og Bullmore, E. (2005). N-back arbeidsminneparadigme: en meta-analyse av normative funksjonelle neuroimaging studier. Hum. Brain Mapp. 25, 46–59. doi: 10.1002 / hbm.20131
PubMed Abstract | CrossRef Full Text/Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S., Og Kumar, S. (2010). Elektrofysiologiske korrelater av kognisjon forbedres med lur under søvnmangel. Eur. J. Appl. Physiol. 108, 549–556. doi: 10.1007 / s00421-009-1222-3
PubMed Abstract | CrossRef Full Text/Google Scholar
Ratcliff, R., Og Van Dongen, H. (2018). Effekten av søvnmangel på element og assosiativ anerkjennelse minne. J. Exp. Psychol. 44, 193–208. doi: 10.1037/xlm0000452
PubMed Abstract | CrossRef Full Text/Google Scholar
Raymond, Ca (1988). Skiftende arbeid, søvnsykluser er på vei til å bli et annet folkehelseproblem. JAMA J. Am. Med. Assoc. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef Full Text / Google Scholar
Saito, H., Yamazaki, H., Matsuoka, H., Matsumoto, K. og Sato, M. (2001). Visuell hendelsesrelatert potensial i mild demens av alzheimers type. Psykiatri Clin. Neurosci. 55, 365–371. doi: 10.1046/j.1440-1819.2001. 00876.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Schacht, A., Werheid, K. Og Sommer, W. (2008). Vurdering av ansikts skjønnhet er rask, men ikke obligatorisk. Cogn. Påvirke. Oppfør deg. Neurosci. 8, 132–142. doi: 10.3758 / cabn.8.2.132
PubMed Abstract | CrossRef Full Text/Google Scholar
Scott, J. P. R., Mcnaughton, L. R., Og Polman, R. C. J. (2006). Effekter av søvnmangel og trening på kognitiv, motorisk ytelse og humør. Physiol. Oppfør deg. 87:408. doi: 10.1016 / j. physbeh.2005.11.009
PubMed Abstract | CrossRef Full Text/Google Scholar
Smallwood, J., Brown, K., Baird, B., Og Schooler, J. W. (2011). Samarbeid mellom standardmodusnettverket og frontal-parietal-nettverket i produksjonen av et internt tankegang. Brain Res. 1428, 60-70. doi: 10.1016 / j.brainres.2011.03.072
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, Me, Mcevoy, Lk, Og Gevins, A. (2002). Virkningen av moderat søvntap på nevrofysiologiske signaler under arbeidsminneoppgaver. Sov 25, 784-794. doi: 10.1093 / sove / 25.7.56
CrossRef Full Text/Google Scholar
Thomas, M. (2003). Nevrale grunnlag for våkenhet og kognitiv funksjonsnedsettelse under søvnighet ii. effekter av 48 og 72 h av søvnmangel på å våkne menneskelig regional hjerneaktivitet. Thalam. Relatert. Syst. 2, 199–229. doi: 10.1016 / S1472-9288 (03) 00020-7
Kryssref Fulltekst | Google Scholar
Av Dongen, H. (2004). Den kumulative kostnaden for ekstra våkenhet: doseresponse effekter på neurobehavioral funksjoner og søvnfysiologi fra kronisk søvnmangel og total søvnmangel. Sov 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef Full Text / Google Scholar
Verweij, I. M., Romeijn, N., Smit, Dj, Og Giovanni, P. (2014). Søvnmangel fører til tap av funksjonell tilkobling i frontale hjernegrupper. BMC Neurosci. 15:88. doi: 10.1186/1471-2202-15-88
PubMed Abstract | CrossRef Full Text/Google Scholar
Wen, X., Liu, Y., Yao, L., Og Ding, M. (2013). Top-down regulering av standardmodus aktivitet i romlig visuell oppmerksomhet. J. Neurosci. 33, 6444–6453. doi: 10.1523 / JNEUROSCI.4939-12. 2013
CrossRef Full Text/Google Scholar
Wiggins, E., Malayka, M. og Kendra, G. (2018). 24-h søvnmangel svekker tidlig oppmerksomhetsmodulasjon av nevralbehandling: en hendelsesrelatert hjernepotensialstudie. Neurosci. Lett. 677, 32–36. doi: 10.1016 / j.neulet.2018.04.022
PubMed Abstract | CrossRef Full Text/Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y., Og Zhou, Q. (2019). Redusert informasjon erstatning av arbeidsminne etter søvnmangel: bevis fra en hendelsesrelatert potensiell studie. Front. Neurosci. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef Full Text/Google Scholar
Zhang, Q., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z., Et al. (2014). En visuell erp-studie av impulshemming etter en zaleplon-indusert lur etter søvnmangel. PLoS En 9: e95653. doi: 10.1371 / tidsskrift.pone.0095653
PubMed Abstrakt | CrossRef Fulltekst/Google Scholar
Leave a Reply