General Microbiology
古細菌は、物理的な類似性のために、もともと細菌であると考えられていた生物のグループです(”archaeabacteria”の最初の名前を説明しています)。 より信頼性の高い遺伝子解析により、古細菌は細菌と真核生物の両方とは異なることが明らかになり、1977年にWoeseによって最初に提案された三つのドメイン分類では、真核生物と細菌と並んで独自のドメインを獲得した。
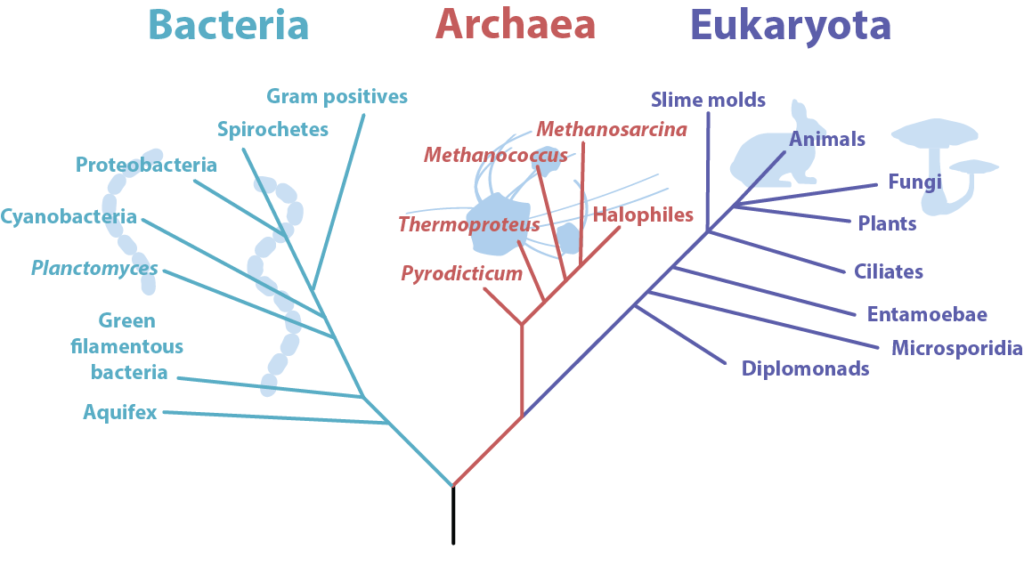
細菌との類似性
では、なぜ古細菌はもともと細菌であると考えられていたのですか? おそらく最も重要なのは、核や他の膜結合細胞小器官がなく、原核生物のカテゴリに入れていることです(伝統的な分類スキームを使用している場合)。 それらのほとんどは単細胞であり、70sサイズのリボソームを有し、典型的には数マイクロメートルの大きさであり、無性生殖のみを行う。 それらは、プラスミド、介在物、鞭毛、および線毛のような、細菌が有することができるのと同じ構造の多くを有することが知られている。 カプセルやスライム層が発見されているが、古細菌ではまれであるように見える。
古細菌はもともと酸、塩、熱の高い場所などの極端な環境から単離されていましたが、”extremophiles”という名前を得ましたが、最近では地表水、海、人間の皮膚、土壌など、細菌が豊富なすべての場所から単離されています。
主な違い
原形質膜
原形質膜にはいくつかの特徴があり、古細菌に特有のものであり、他のドメインとは異なります。 そのような特徴の一つは、ホスホリピドヘッドと側鎖との間のグリセロール結合のキラリティーである。 古細菌では、それはL-異性体の形態であり、細菌および真核生物はD-異性体の形態を有する。 第二の違いは、細菌および真核生物に見られるエステル結合脂質とは対照的に、グリセロールと側鎖との間のエーテル結合の存在である。 エーテル結合は、膜により多くの化学的安定性を提供する。 イソプレノイド鎖は古細菌で発見されている間、第三および第四の違いは、側鎖自体、細菌および真核生物の非分岐脂肪酸に関連付けられています。 これらのイソプレノイド鎖は、分岐側鎖を有することができる。細菌と古細菌の間の原形質膜脂質の比較。
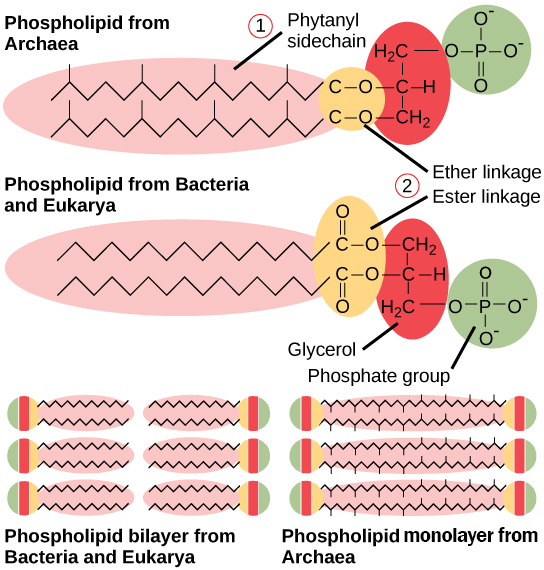
最後に、古細菌の原形質膜は、一つのリン脂質のイソプレン鎖が膜の反対側のリン脂質のイソプレン鎖と接続する単分子層として見 細菌と真核生物は脂質二重層のみを有し、膜の両側は分離されたままである。細菌と同様に、古細菌の細胞壁は、環境および細胞内の圧力から細胞を保護するように設計された半剛性構造である。
細胞壁
細菌の細胞壁は典型的にはペプチドグリカンを含むが、その特定の化学物質は古細菌には欠けている。 代わりに、古細菌は、生物の環境に適応した細胞壁の種類の多種多様を表示します。 いくつかの古細菌は完全に細胞壁を欠いています。
それは普遍的ではありませんが、多数の古細菌は、細胞壁自体の一部であると考えられるタンパク質性のS層を持っています(細菌とは異なり、S層は細胞壁に加えて構造である)。 いくつかの古細菌では、S層は唯一の細胞壁成分であり、他のものでは追加の成分によって結合されている(下記参照)。 古細菌のS層は、多くの場合、細胞の原形質膜に固定され、タンパク質または糖タンパク質のいずれかで作ることができます。 蛋白質は滑らかな外の表面が付いている二次元の結晶の配列を形作ります。 いくつかのS層は、二つの異なるS層タンパク質から構成されています。
古細菌はペプチドグリカンを欠いていますが、いくつかはpseudomureinとして知られている同様の化学構造を持つ物質を含んでいます。 NAMの代りに、それは強さを高めるためにペプチッドinterbridgesが付いているN-acetylalosaminuronic酸(NAT)をNAGに、つながりました含んでいます。
メタノコンドロイチンは、脊椎動物に見られる結合組織成分コンドロイチンと組成が類似しているいくつかの古細菌細胞に見られる細胞壁ポ
いくつかの古細菌は、S層に類似した格子構造で構成されるタンパク質鞘を有する。 しかし、これらの細胞はしばしば糸状鎖に見られ、個々の細胞とは対照的に、タンパク質鞘は鎖全体を取り囲んでいる。/p>
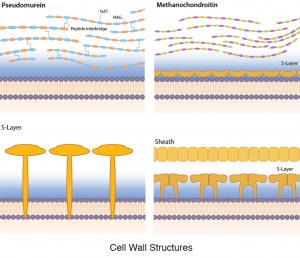
細胞壁の構造的多様性。
リボソーム
古細菌は細菌と同じサイズの70Sのリボソームを持っていますが、rrnaのヌクレオチドの違いが、古細菌は細菌とは別のドメインに値すると主張する決定的な証拠を科学者に提供しました。 さらに、古細菌のリボソームは、細菌のリボソームとは異なる形状を有し、古細菌に特有のタンパク質を有する。 これは細菌のribosomal機能を禁じる抗生物質への抵抗をそれらに与えます。
構造
細菌に見られる構造の多くは古細菌でも発見されていますが、それぞれの構造が物質と構造の違いに基づいて独立して進化した
Cannulae
cannulae、古細菌に固有の構造は、いくつかの海洋古細菌株で発見されています。 これらの中空の管のような構造は、分裂後に細胞を接続するように見え、最終的には多数の細胞と管で構成される密なネットワークにつながる。 これは、細胞のコミュニティを表面に固定する手段として役立つ可能性があります。
ハムス(pl. hami)
古細菌に固有のもう一つの構造は、hamus、遠端に三つのフックを持つ長い螺旋管です。 ハミは、細胞が互いに表面の両方に付着することを可能にし、コミュニティの形成を促進するように見える。
Pilus(pl. pili)
Piliは古細菌で観察されており、細菌のpilinから改変された可能性が最も高いタンパク質で構成されています。 得られた管様構造は、表面への付着に使用されることが示されている。
鞭毛(pl. 鞭毛)
古細菌の鞭毛は、運動性に使用されているが、細菌の鞭毛とは著しく異なるため、細菌の対応物と区別するために”古細菌”と呼ぶことが提案されている。
細菌の鞭毛と古細菌の鞭毛の間には何が似ていますか? 両方とも細胞が細胞から伸びる堅いフィラメントの回転によって推進される動きのために使用される。 その後、類似点は終了します。違いは何ですか?
古細菌の鞭毛の回転は、細菌で使用される陽子の原動力とは対照的に、ATPによって駆動される。 古細菌の鞭毛を構成する蛋白質は細菌の鞭毛よりもむしろ細菌のpiliで、見つけられる蛋白質に類似しています。 古細菌の鞭毛フィラメントは中空ではないので、鞭毛タンパク質が末端に添加されるのではなく、フィラメントの基部に挿入されると成長が起こる。 フィラメントはいくつかの異なるタイプのフラジェリンで構成されていますが、細菌の鞭毛フィラメントには1つのタイプが使用されます。 時計回りの回転は古細菌細胞を前方に押し、反時計回りの回転は古細菌細胞を後方に引っ張ります。 ランとタンブリングの交代は観察されません。
分類
現在、古細菌にはEuryarchaeotaとProteoarchaeotaの二つの認識された門があります。 いくつかの追加の門が提案されている(Nanoarchaeota、Korarchaeota、Aigarchaeota、Lokiarchaeota)が、主に証拠が環境配列のみから来ているという事実のために、公式にはまだ認識されていない。
キーワード
古細菌、L-異性体型、D-異性体型、エーテル結合、エステル結合、イソプレノイド鎖、分岐側鎖、脂質単分子層、脂質二重層、S層、プソードムレイン、N-アセチルアロサミヌロン酸(NAT)、メタノコンドロイチン、タンパク質鞘、カニューレ、hamus/hami、pilus/pili、鞭毛/鞭毛、古細菌、euryarchaeota、Proteoarchaeota。古細菌はどのように細菌に似ていますか?
必須の質問/目的
- 古細菌はどのように細菌に似ていますか?
- 細菌と比較して、古細菌の原形質膜の違いを説明します&真核生物。 違いを説明します。
- 古細菌にはどのような種類の細胞壁が存在し、それらは何から構成されていますか?
- 古細菌のリボソームはどのように類似しており、細菌のリボソームとは異なるのですか?
- 古細菌の線毛は細菌の線毛とどのように違うのですか?
- カニューレとハミとは何ですか? 彼らは古細菌のためにどのような役割を果たすことができますか?
- 古細菌の鞭毛は、組成、組み立て、および機能の点で、細菌の鞭毛とどのように異なるのですか?
- 物理的特性の面で、古細菌と細菌の共通点と違いを理解しています。古細菌は細菌と物理的に類似しているにもかかわらず、真核生物とより密接に関連しているように見えるという事実を説明するものは何ですか?
Leave a Reply