ペプチド結合
窒素原子上の孤立電子対の重要な非局在化は、グループに部分的な二重結合の特性を与える。 部分的な二重結合はアミド基を平面にし、シス異性体またはトランス異性体のいずれかに生じる。 タンパク質の展開状態では、ペプチド基は自由に異性化し、両方の異性体を採用するが、折り畳まれた状態では、各位置に単一の異性体のみが採用される(まれな例外を除く)。 トランス形態は、ほとんどのペプチド結合において圧倒的に好ましい(トランス:シス集団における約1000:1の比)。 しかし、X-Proペプチド群は、おそらくC α{\displaystyle\mathrm{C^{\alpha}}}

とC δ{\displaystyle\mathrm{C^{\delta}}}

プロリンの原子は、cis異性体とtrans異性体をエネルギーでほぼ等しくします(下の図を参照)。
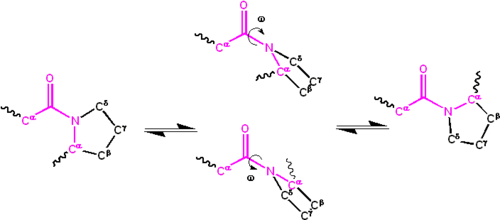
ペプチド群に関連する二面角(四つの原子C α−C’-N−C α{\displaystyle C^{\alpha}-C^{\prime}-N−C^{\alpha}}で定義される)
 )はω{\displaystyle\omega}
)はω{\displaystyle\omega}

と表される。 ; ω=0∞{\displaystyle\omega=0^{\circ}}

シス異性体(シンペリプラナー立体配座)およびω=180∞{\displaystyle\omega=180^{\circ}}
 トランス異性体(antiperiplanar立体配座)のために。 アミド基はcis型とtrans型の間のC’-N結合の周りで異性化することができるが、ゆっくりと異性化することができる(室温で20秒)。 遷移状態ω=±90π{\displaystyle\omega=\pm90^{\circ}}
トランス異性体(antiperiplanar立体配座)のために。 アミド基はcis型とtrans型の間のC’-N結合の周りで異性化することができるが、ゆっくりと異性化することができる(室温で20秒)。 遷移状態ω=±90π{\displaystyle\omega=\pm90^{\circ}}

は部分的な二重結合を破壊する必要があるため、活性化エネルギーはおよそ80キロジュール/mol(20kcal/mol)である。 しかし、活性化エネルギーは、疎水性環境にペプチド基を配置したり、x-Proペプチド基の窒素原子に水素結合を供与するなど、単一結合形態を好む変化によ 活性化エネルギーを低下させるためのこれらのメカニズムの両方が、X-Proペプチド結合のシス-トランス異性化を触媒する天然に存在する酵素であるペプチジルプロリルイソメラーゼ(Ppiase)で観察されている。
立体配座タンパク質の折り畳みは、通常、cis-trans異性化(10-100s)よりもはるかに高速(通常は10-100ms)です。 あるペプチッドグループのnonnative異性体はそれを減速するか、または天然異性体が達されるまで起こることからそれを防ぐconformational折りたたみをかなり破壊でき しかし、すべてのペプチド基が折り畳みに同じ効果を有するわけではない; 他のペプチド基の非結合異性体は、折り畳みに全く影響しない可能性がある。
Leave a Reply