Liaison peptidique
Une délocalisation significative de la seule paire d’électrons sur l’atome d’azote confère au groupe un caractère de double liaison partielle. La double liaison partielle rend le groupe amide planaire, se produisant dans les isomères cis ou trans. À l’état déplié des protéines, les groupes peptidiques sont libres de s’isomériser et d’adopter les deux isomères ; cependant, à l’état replié, un seul isomère est adopté à chaque position (à de rares exceptions près). La forme trans est préférée massivement dans la plupart des liaisons peptidiques (rapport d’environ 1000: 1 dans les populations trans: cis). Cependant, les groupes peptidiques X-Pro ont tendance à avoir un rapport d’environ 30: 1, probablement parce que la symétrie entre le C α {\displaystyle\mathrm {C^{\alpha}}}

et le C δ {\displaystyle\mathrm {C^{\delta}}}

les atomes de proline rendent les isomères cis et trans presque égaux en énergie (voir figure ci-dessous).
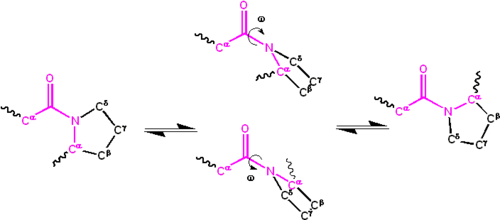
L’angle dièdre associé au groupe peptidique (défini par les quatre atomes C α−C’−N−C α {\displaystyle C ^{\alpha}- C^{\prime} – N-C^{\alpha}}

) est noté ω {\displaystyle\omega}

; ω=0 { {\displaystyle\omega= 0^{\circ}}

pour l’isomère cis (conformation synperiplanaire) et ω=180 {{\displaystyle\omega=180^{\circ}}

pour l’isomère trans (conformation antipériplanaire). Les groupes amides peuvent isomériser sur la liaison C’-N entre les formes cis et trans, bien que lentement (τ ∼{\displaystyle\tau\sim}

20 secondes à température ambiante). Les états de transition ω = ± 90 {{\displaystyle\omega =\pm 90^{\circ}}

nécessite que la double liaison partielle soit rompue, de sorte que l’énergie d’activation soit d’environ 80 kilojoule/mol (20 kcal/mol). Cependant, l’énergie d’activation peut être réduite (et l’isomérisation catalysée) par des changements qui favorisent la forme monocollée, tels que le placement du groupe peptidique dans un environnement hydrophobe ou le don d’une liaison hydrogène à l’atome d’azote d’un groupe peptidique X-Pro. Ces deux mécanismes d’abaissement de l’énergie d’activation ont été observés dans les peptidyl prolyl isomérases (PPIases), qui sont des enzymes naturelles qui catalysent l’isomérisation cis-trans des liaisons peptidiques X-Pro.
Le repliement conformationnel des protéines est généralement beaucoup plus rapide (typiquement 10-100 ms) que l’isomérisation cis-trans (10-100 s). Un isomère non nonnatif de certains groupes peptidiques peut perturber le repliement conformationnel de manière significative, soit en le ralentissant, soit en l’empêchant de se produire jusqu’à ce que l’isomère natif soit atteint. Cependant, tous les groupes peptidiques n’ont pas le même effet sur le pliage; les isomères non natifs d’autres groupes peptidiques peuvent ne pas affecter du tout le repliement.
Leave a Reply