Frontières en Neurosciences
Introduction
Avec les progrès de la société et les changements de rythme de travail, un nombre croissant de personnes souffrent de privation de sommeil. La privation de sommeil nuit non seulement à la santé physique et mentale de l’individu, mais affecte également gravement la performance au travail, provoquant des erreurs de travail et même des accidents. Par conséquent, comprendre le mécanisme de la privation de sommeil qui affecte la fonction cognitive est d’une grande importance pour prévenir efficacement les effets de la privation de sommeil.
Des études antérieures ont révélé que la privation de sommeil peut entraîner une série de changements dans l’humeur, la capacité cognitive, le rendement au travail et la fonction immunitaire d’un individu (Choo et al., 2005). Le manque de sommeil perturbe la circulation corporelle et affecte les capacités cognitives et émotionnelles des individus (Raymond, 1988). Plusieurs études ont révélé que la privation de sommeil nuit à l’inhibition de la réponse (Harrison et Horne, 1998; Muzur et al., 2002; Jennings et coll., 2003). Par exemple, après 36 h de privation de sommeil, la capacité de l’individu à supprimer les stimuli négatifs a diminué (Chuah et al., 2006). Des études de neuroimagerie ont suggéré que la privation de sommeil réduit le faible niveau de capacité de traitement visuel d’un individu (Anderson et Platten, 2011; Ning et al., 2014). De plus, la privation de sommeil altère l’hippocampe et pourrait affecter la mémoire en détruisant la plasticité synaptique (Cote et al., 2014). Thomas (2003) a indiqué que le manque de sommeil réduisait le flux sanguin cérébral et le taux métabolique dans le thalamus, le cortex préfrontal et le cortex pariétal (Géraldine et al., 2005). Jarraya et ses collègues ont constaté que la privation partielle de sommeil affectait de manière significative les fonctions neuropsychologiques telles que la mémoire instantanée verbale, l’attention et la vigilance (Thomas, 2003). De plus, certaines études ont révélé que les effets cumulatifs d’une privation partielle de sommeil pourraient gravement altérer les fonctions cognitives et le comportement (Van Dongen, 2004; Scott et al., 2006; Jarraya et coll., 2013).
La mémoire de travail est un système utilisé pour stocker et traiter l’information et qui est une fonction cognitive à capacité limitée (Bartel et al., 2004). De plus, les informations stockées dans le système de mémoire de travail peuvent être modifiées de la mémoire à court terme à la mémoire à long terme par le biais de la répétition et d’autres méthodes de mémoire. La mémoire de travail est la transition entre les systèmes de mémoire à court et à long terme, ce qui est très essentiel dans le traitement des messages humains (Miyake et Shah, 1999). Il fournit un espace de stockage temporaire et les ressources nécessaires pour traiter les informations, telles que la compréhension vocale, le raisonnement et l’apprentissage. Il a été démontré que la privation de sommeil affecte d’abord la mémoire de travail.
Des études antérieures ont utilisé le paradigme de la mémoire de travail n-back chez des participants qui ont subi une privation de sommeil et ont constaté que le manque de sommeil induit une diminution de l’activité métabolique dans le réseau régional du cerveau, principalement le traitement de l’information et l’inhibition de la réaction (Baddeley, 2000; Zhang et al., 2019). Une altération de la mémoire de travail après une privation de sommeil est liée à l’activation du réseau par défaut dans les tâches (Chee et Chuah, 2008), ce qui peut être lié au rôle important du thalamus dans la vigilance corticale. Par exemple, la privation de sommeil augmentait la connexion entre l’hippocampe, le thalamus et le réseau par défaut, ce qui s’accompagnait souvent d’une somnolence subjective plus élevée et d’une moins bonne performance de la mémoire de travail (Lei et al., 2015; Li et coll., 2016). Études sur la privation de sommeil identi?l’augmentation de la latence et la réduction de l’amplitude de la composante P3 étaient associées à une sobriété prolongée (Morris et al., 1992; Jones et Harrison, 2001; Panjwani et coll., 2010). La diminution de l’onde P3 pourrait refléter une diminution de l’attention des participants et une réduction du discernement des stimuli cibles (Koslowsky et Babkoff, 1992).
Cependant, peu d’études ont fourni des preuves électrophysiologiques pour une altération de la mémoire de travail après une privation de sommeil. La tâche n-back est considérée comme une méthode courante pour évaluer la mémoire de travail (Owen et al., 2005; Jaeggi et coll., 2010). Zhang et coll. conçu une tâche de mémoire de travail de prononciation à deux dos pour explorer l’alternance de messages diminués de la mémoire de travail pendant la privation de sommeil, mais peu d’études ont utilisé différents types de tâches de mémoire de travail dans une seule expérience. Dans la présente étude, nous avons conçu différents types de tâches de mémoire de travail (mémoire de travail de prononciation, mémoire de travail spatiale et mémoire de travail d’objet) pour explorer l’altération de la fonction cognitive par TSD et les données EEG des participants enregistrées à 2 moments (ligne de base et 36 h-TSD). Toutes les tâches ont adopté un paradigme à 2 dos. Cette étude a évalué les changements de l’onde N2-P3 liés à la mémoire de travail pendant le TSD et a analysé les caractéristiques temporelles des effets de la privation de sommeil sur la mémoire de travail. Nos résultats fournissent des preuves expérimentales des effets de la privation de sommeil sur la fonction cognitive.
Matériaux et méthodes
Participants
Seize jeunes étudiants droitiers en bonne santé ont participé à cette étude. Nous avons recruté des participants par la publicité sur le campus. Les participants avaient tous de bonnes habitudes de sommeil (PSQI<5). Tous les participants étaient âgés de 21 à 28 ans avec une moyenne d’âge de 23 ans, et aucun des participants n’avait de maladie mentale ou physique. Tous les participants avaient une vision normale ou une vision corrigée supérieure à 1,0 et des scores d’intelligence > 110 au test Raven. Avant l’expérience, l’expérimentateur a expliqué la procédure et les points d’attention aux participants pour s’assurer qu’ils connaissaient bien la méthode et la procédure. Au cours des 2 semaines précédant l’expérience, les participants dormaient régulièrement pendant 7 à 9 h par jour, sans fumer, boire du café, boire de l’alcool ou consommer de médicaments pendant 2 jours avant l’expérience. Avant l’expérience, tous les participants ont fourni un consentement éclairé écrit. Le schéma expérimental a été approuvé par le Comité d’éthique de la Quatrième Université de médecine militaire et de l’Université de Beihang.
Conception expérimentale
Trois types de tâches de mémoire de travail ont été présentés à tous les participants. Il s’agissait d’une tâche de mémoire de travail de prononciation à deux dos (voir Figure 1), d’une tâche de mémoire de travail spatiale à deux dos (voir Figure 2) et d’une tâche de mémoire de travail d’objet à deux dos (voir Figure 3). Les matériaux stimulants des tâches étaient 15 lettres anglaises insensibles à la casse qui excluaient celles avec des lettres similaires, telles que L / l, M / m; petits carrés noirs; et 12 figures géométriques, respectivement. Tous les matériaux ont été présentés en couleur noire sur fond blanc, avec un angle visuel approximatif de 1,5 ° × 1,5 ° (largeur: 2,0 cm, hauteur: 2,0 cm). 122 pistes ont été comprises dans chaque tâche et, dans chaque piste, le stimulus cible a été présenté pendant 400 ms deux pistes après la présentation du stimulus objectif, avec le temps d’asynchronie d’apparition du stimulus de 1 600 ms (SOA) qui a été marqué par un « + blanc.”Les participants ont été invités à cliquer sur le bouton gauche de la souris lorsque la cible et le stimulus objectif étaient identiques (« correspondance”), tandis que cliquez sur le bouton droit de la souris lorsqu’ils ne l’étaient pas (« inadéquation”). La condition correspondante ou non a été présentée dans un ordre pseudo-aléatoire avec un rapport de 1: 1.
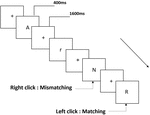
Figure 1. Schéma de la tâche de mémoire de travail de prononciation.
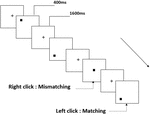
Figure 2. Schéma de la tâche de mémoire de travail spatiale.
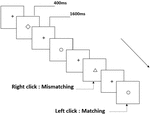
Figure 3. Schéma de la tâche mémoire de travail de l’objet.
Procédures expérimentales
Avant l’expérience, les participants ont été informés de la tâche expérimentale. Ils ont été informés de la pratique des trois types de tâches de mémoire de travail jusqu’à ce qu’un taux de précision de 90% soit atteint. Les participants ont visité le laboratoire la veille de l’expérience et ont dormi dans le laboratoire cette nuit-là. Les deux participants partenaires ont effectué les expériences en même temps. Trois types de tâches de mémoire de travail ont été effectuées de 7h30 à 8h30 le lendemain matin avec enregistrement simultané de l’électroencéphalogramme (EEG) (ligne de base). Le deuxième enregistrement EEG (36 h-TSD) a été effectué après une période de 36 h pendant laquelle les participants n’ont pas été autorisés à dormir. Pendant toute la durée de l’expérience, l’inhibition centrale et les médicaments stimulants ont été interdits. Les participants ont été accompagnés, observés et rappelés par le personnel infirmier afin de les garder éveillés tout au long de la session TSD.
Enregistrements EEG
Un EEG scalp continu a été enregistré à l’aide de capuchons d’électrodes placés à 64 endroits à l’aide du système 10-20 avec un amplificateur SynAmps2. Les mastoïdes bilatéraux (A1 et A2) ont été utilisés à titre de référence et le front a été utilisé comme sol. Les EEG ont été enregistrés à 1 000 Hz et l’impédance de tous les canaux a été maintenue en dessous de 5 kΩ. Quatre électrodes supplémentaires ont été placées au-dessus et au-dessous des yeux droit et gauche pour enregistrer un électrooculogramme bipolaire vertical et horizontal.
Analyse des données des expériences comportementales
En raison d’erreurs techniques, deux cas ont été supprimés tandis que 14 autres cas ont été inclus dans l’analyse statistique suivante. Les données comportementales comprenaient le temps de réaction moyen, le taux correct et le nombre correct par unité de temps. Les données comportementales dans les états de base et 36 h-TSD ont été enregistrées pour analyse. Les analyses ont été effectuées par IBM SPSS (V22.2), où la méthode d’analyse de variance par mesures répétées (ANOVA) avec Greenhouse-Geisser était une analyse post-hoc de Bonferroni a été lancée. Les résultats statistiques ont été présentés sous la forme de la moyenne et de l’écart type (ET).
Analyse des données EEG
Le programme Scan 4.3 a été utilisé pour analyser les données EEG, où les artefacts EEG du mouvement oculaire ont été corrigés par la méthode de réduction des artefacts oculaires. Des époques allant de -100 à 800 ms des données EEG continues ont été extraites et filtrées par un filtre passe-bande de 0,5 à 30 Hz avec une pente de fréquence de 24 dB/oct. Les essais dans lesquels la tension dépassait ± 100 µV ont été rejetés et la ligne de base a été corrigée à une amplitude moyenne de 100 ms. Les composantes de l’EEG ont été moyennées et calculées avec uniquement les réponses corrigées. Les composants ERP P2 (100-250 ms), N2 (150-350 ms) et P3 (250-450 ms) des essais de stimulus ont été identifiés et quantifiés. Les amplitudes de crête et les latences grand-moyennes des composantes N2 et P3 ont été calculées séparément à F3, Fz, F4, C3, Cz, C4, P3, Pz et P4, et la composante P2 a été calculée à F3, Fz, F4, C3, Cz et C4 (Casement et al., 2006; Verweij et coll., 2014).
Des mesures répétées de l’ANOVA ont été utilisées pour tous les résultats de l’ERP. Les principaux effets et les interactions entre les états de sommeil (ligne de base et 36 h-TSD), les tâches (mémoire de travail de prononciation, mémoire de travail spatiale et mémoire de travail d’objet), les régions (frontales, centrales et pariétales; la composante P2 a été analysée uniquement sur les régions frontales et centrales) et les sites (gauche, milieu et droite) ont été analysés statistiquement en utilisant des mesures répétées ANOVA, qui incluaient des corrections de Greenhouse-Geisser pour la non-sphéricité et des tests post-hoc de Bonferroni.
Résultats
Performance comportementale
Les résultats des expériences comportementales sont présentés dans le tableau 1. Le temps de réaction moyen était plus long à l’état de 36 h-TSD qu’au départ avec une tendance à l’augmentation mais sans différences significatives. ANOVA a révélé que le taux correct de la tâche était significativement différent entre la ligne de base et 36 h-TSD. Le nombre correct par unité de temps a montré un effet principal significatif du temps pendant 36 h-TSD.

Tableau 1. Données de performance (moyenne ± écart-type) sur la tâche 2-back au départ et après 36 h-TSD.
Amplitude
Par rapport à la ligne de base, une diminution significative de l’amplitude de P3 a été observée et une augmentation significative de l’amplitude de P2 après TSD. Bien que l’amplitude de N2 ait diminué après 36 h de TSD, la différence n’a pas atteint de signification statistique (tableau 2).

Tableau 2. Amplitude maximale moyenne des composantes P2, N2 et P3 dans la condition de réponse correcte sur plusieurs sites d’électrodes à la ligne de base et après 36 h-TSD.
Des effets principaux significatifs des régions et des sites sur l’amplitude P2 ont été trouvés dans la condition TSD. Pendant le TSD, l’amplitude maximale de P2 est apparue dans la région frontale (Figure 4). De plus, les différences d’amplitudes P2 dans les différentes régions (frontale vs centrale) étaient plus significatives dans les électrodes bilatérales (à gauche: P = 0,001; à droite: P = 0,000) que dans les électrodes du milieu (Figure 4). Un effet principal significatif de la région et du site sur N2 a révélé que l’amplitude de N2 était plus négative dans la région frontale que dans la région centrale (P = 0,008, Figure 4) et était plus petite à droite que sur le côté gauche (P = 0,011, Figures 5A2, B2). Un effet principal du site sur l’amplitude P3 a été observé. L’amplitude de P3 était plus positive au milieu que sur le côté gauche (P = 0,009, Figure 4). Un effet d’interaction significatif entre le temps et la région a été observé pour l’amplitude P3. Pendant le TSD, la réduction de l’amplitude de P3 était plus significative dans les régions frontales et centrales que dans la région pariétale (P = 0,005; P = 0,003) (Figure 5C3). Aucun autre effet principal ou effet d’interaction n’a atteint une signification statistique.
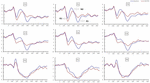
Figure 4. Amplitude ERP à la ligne de base et 36 h-TSD pour la condition de réponse correcte pour la tâche de mémoire de travail. Les canaux sont ordonnés de gauche à droite et de haut en bas comme suit: F3, Fz, F4, C3, Cz, C4, P3, Pz et P4. Par rapport à la ligne de base, les latences des composantes N2-P3 ont été prolongées et les amplitudes de N2-P3 ont été diminuées après 36 h-TSD.
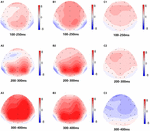
Figure 5. Carte topographique de la réponse correcte dans la tâche de mémoire de travail dans différentes conditions de sommeil (A1-C3). (A1) P2, 100-250 ms, au départ. (A2) N2, 200-300 ms, au départ. (A3) P3, 300-400 ms, au niveau de référence. (B1) P2, 100-250 ms, à 36 h – TSD. (B2) N2, 200-300 ms, pour 36 h-TSD. (B3) P3, 300-400 ms, pour 36 h-TSD. (C1) P2, 100-250 ms, 36 h-TSD avec ligne de base soustraite. (C2) N2, 200-300 ms, 36 h-TSD avec référence soustraite. (C3) P3, 300-400 ms, 36 h-TSD avec référence soustraite.
Latence
Les latences de N2 et de P2 ont été significativement prolongées après TSD. Bien que la latence P3 ait été prolongée après 36 h de TSD, la différence n’a pas atteint de signification statistique (tableau 3).

Tableau 3. Latence maximale moyenne des composants P2, N2 et P3 dans la condition de réponse correcte sur plusieurs sites d’électrodes à la ligne de base et après 36 h-TSD.
L’effet principal significatif de la région sur N2 et P3 a révélé que la latence des composantes N2-P3 était plus courte dans la région pariétale que dans la région frontale (P = 0,002; P = 0,000) et la région centrale (P = 0,000; P = 0,000) (Figure 4). La latence de l’onde P3 était significativement plus longue du côté gauche que du côté droit (Figure 4).
Aucun autre effet principal ou effet d’interaction n’a atteint une signification statistique.
Les amplitudes et latences N2, P2 et P3 qui ont été déclenchées aux neuf sites d’électrodes sont présentées à la figure 4. La carte topographique de la réponse correcte dans la tâche de mémoire de travail dans différentes conditions de sommeil (ligne de base, 36 h-TSD et la différence entre les deux conditions) est présentée à la figure 5.
Discussion
Dans cette étude, nous avons rapporté les influences de la privation de sommeil de 36 h sur la mémoire de travail, en combinant des données comportementales dans deux états de sommeil (ligne de base et 36 h-TSD) avec des enregistrements EEG contemporains. L’analyse des résultats a indiqué que les changements dans les données comportementales en fonction de la mémoire de travail altérée après 36 h TSD: une augmentation du temps de réaction moyen des tâches cognitives et une diminution de la précision.
La privation de sommeil a altéré le contrôle des ressources attentionnelles de l’individu. Bien que les individus aient essayé de maintenir l’éveil et la performance au travail, y compris le temps de réaction et le taux correct, pendant la privation de sommeil, la capacité de traitement de l’information de leur mémoire de travail était toujours affectée en raison de la diminution de la vitesse de traitement de l’information (Casement et al., 2006; Wiggins et coll., 2018). Dans cette étude, les ondes N2 et P3 liées à la mémoire de travail ont été mesurées pour montrer une augmentation de la latence et une diminution de l’amplitude après une privation de sommeil par rapport aux lectures de base. Des études ont démontré que la privation de sommeil entraîne une baisse continue de l’attention, et le phénomène de diminution de l’amplitude P3 indique que le contrôle descendant de la cognition des individus s’effondre progressivement. La privation de sommeil a un effet plus néfaste sur les fonctions cognitives, en particulier celles qui dépendent des fonctions mentales ou cognitives (Kusztor et al., 2019).
La composante P3 reflète le déploiement des ressources d’attention, et la latence de P3 est largement considérée comme la fenêtre temporelle de catégorisation et d’évaluation des stimulus. La diminution de l’amplitude de l’onde P3 a également confirmé que la prise de décision dans la réponse d’appariement après que le DNT ait été endommagé dans une certaine mesure (Gosselin et al., 2005). Des études ont suggéré que la privation de sommeil peut affecter l’étape de traitement de l’information de la mémoire de travail. Dans cette étude, les indicateurs de performance ont également soutenu la conclusion que le temps de réponse au stimulus cible était augmenté et que la latence de l’onde P3 était prolongée (Cote et al., 2008). Il a été émis l’hypothèse que l’effet de la privation de sommeil sur les composants P3 pourrait également avoir lieu en raison de l’incapacité de répondre à l’altération de l’information, ce qui est cohérent avec les conclusions précédentes selon lesquelles les composants P3 sont liés à la mise à jour du contenu de la mémoire de travail (Donchin et Fabiani, 1991).
Des études antérieures ont considéré le composant N2 comme un indice électrophysiologique reflétant la capacité de l’individu à supprimer la réponse (Kreusch et al., 2014). Après une privation de sommeil, la latence prolongée de la composante NoGo-N2 indique que la capacité de l’individu à supprimer la réponse est altérée (Jin et al., 2015). La diminution de l’amplitude et la latence prolongée de la composante N2 liée à la mémoire de travail de prononciation après la privation de sommeil révèlent que la privation de sommeil nuit au traitement de l’information de la mémoire de travail de prononciation (Zhang et al., 2019). On pense généralement que la composante N2 reflète l’attention sélective du cerveau et le traitement des stimuli ou des signaux émotionnels (Schacht et al., 2008) et est une composante endogène liée à l’état mental, à l’attention et au degré d’attention d’un individu. Dans cette étude, nous avons constaté que la latence de la composante N2 était significativement prolongée, mais que l’amplitude ne montrait qu’une tendance à la baisse. Selon des études antérieures, une latence prolongée de N2 reflétait une augmentation du temps de réponse après la restriction du sommeil (Zhang et al., 2014). Cependant, la conclusion selon laquelle l’amplitude de N2 n’a pas été modifiée de manière significative peut être due à des réponses compensatoires cérébrales (Drummond et Brown, 2001). Dans le cas de ressources cognitives limitées, il existait un mécanisme de compensation pour restaurer la fonction cognitive altérée (Jin et al., 2015).
Selon la topographie du cuir chevelu, les changements des composants N2-P3 liés à la privation de sommeil sont plus évidents dans la zone frontale. Le contrôle frontopariétal (FPC) joue un rôle important dans le contrôle cognitif. Des études ont montré que le CPF peut contourner le contrôle cognitif descendant, permettant aux individus de se concentrer sur les informations liées à la cible tout en supprimant les informations qui ne sont pas liées à la cible (Smallwood et al., 2011; Wen et coll., 2013). Le CPF est important pour la conservation et le traitement de l’information dans la mémoire de travail, et le degré d’activation du CPF après la privation de sommeil a été réduit par rapport à un groupe de sommeil normal (Ma et al., 2014). Bien que les résultats de l’EEG ne reflètent pas en détail les changements dans des régions spécifiques du cerveau, ils reflètent intuitivement l’effet du TSD sur la rétention et le traitement des informations de la mémoire de travail.
Bien que le processus cognitif exact sous-jacent à la composante P2 soit encore largement débattu, en tant que définition large, la composante P2 reflète le processus d’attention et de traitement visuel et est généralement considérée comme liée à l’attention sélective et à la mémoire de travail, reflétant le jugement précoce du processus perceptuel (Saito et al., 2001). Dans cette étude, nous avons constaté une augmentation significative de l’amplitude de l’onde P2 après une privation de sommeil. Des études ont rapporté que les ondes P2 pourraient faire partie du système d’appariement cognitif précoce pour le traitement des messages et peuvent comparer les entrées sensorielles à la mémoire stockée (Freunberger et al., 2007) sont sensibles aux altérations de l’attention de la mission et des exigences de mémoire de travail (Smith et al., 2002). La compensation fonctionnelle est l’une des fonctions uniques du cerveau humain et un facteur important pour le maintien de la fonction cognitive. Une activation excessive du cortex préfrontal dorsolatéral (DLPFC) après une privation de sommeil indique que, à mesure que les ressources cérébrales diminuent, le DLPFC semble avoir une fonction compensatoire (Drummond et al., 2004; Choo et coll., 2005). Par conséquent, nous supposons que l’augmentation significative de l’amplitude P2 observée dans cette étude peut être due à une compensation fonctionnelle dans laquelle les individus semblent maintenir une fonction cognitive normale après une privation de sommeil. Bien qu’un grand nombre d’études aient utilisé la technologie ERP pour explorer l’effet de la privation de sommeil sur les fonctions cognitives, les composants précoces tels que N1 et P2 n’ont pas été systématiquement étudiés et les résultats sont incohérents (Evans et Federmeier, 2007; Wiggins et al., 2018; Zhang et coll., 2019). Peu de recherches explorent le changement de la composante P2 pendant la privation de sommeil (Mograss et al., 2009). Par conséquent, les effets de la privation de sommeil sur les premiers composants de l’ERP, tels que P2, doivent encore être étudiés et explorés.
Dans cette expérience, nous avons utilisé le modèle 2-back pour concevoir des tâches de mémoire de travail de prononciation, d’espace et d’objet et avons examiné l’altération de la mémoire de travail après 36 h de TSD. Par rapport aux études précédentes qui se concentraient uniquement sur les effets de la privation de sommeil sur un type spécifique d’informations, telles que la mémoire de travail de prononciation, ou une fonction cognitive spécifique, telle que l’inhibition de la réponse, nous avons examiné le contenu du modèle de mémoire de travail et analysé de manière exhaustive les effets de la privation de sommeil sur la mémoire de travail.
Cependant, l’étude présente certaines limites. Tout d’abord, nous n’avons utilisé que la tâche 2-back et n’avons pas comparé les performances des participants dans des tâches de mémoire de travail de différentes difficultés. Par conséquent, il existe des limites pour expliquer et inférer les changements dans la charge de travail. Deuxièmement, seuls des volontaires de sexe masculin ont été utilisés dans l’étude, et les conclusions doivent être évaluées lors de leur extension aux volontaires de sexe féminin. En raison du nombre limité de participants, nous avons seulement constaté que l’amplitude de l’onde N2 avait une tendance à la baisse et que la latence de l’onde P3 avait une tendance prolongée. Des résultats stables pourraient être obtenus après l’augmentation du nombre de participants. Troisièmement, la combinaison de notre procédure avec l’IRMf pour la mémoire de travail peut faciliter une interprétation ultérieure des résultats. Des études antérieures ont montré que les biorythmes circadiens affectent les performances comportementales et qu’il existe des différences individuelles (Montplaisir, 1981; Lavie, 2001). Nous n’avons pas enregistré les données EEG au même moment dans cette expérience, de sorte que l’influence des biorythmes circadiens sur les résultats des tests ne peut pas être complètement exclue.
Cette recherche a montré que la capacité de mémoire de travail était altérée après TSD et que ces dommages n’étaient pas associés au contenu de stimulus de la mémoire de travail. Le manque de sommeil a réduit la qualité des informations stockées en mémoire, ce qui pourrait se produire avec le processus dégénératif de l’attention (Ratcliff et Van Dongen, 2018). Cette étude fournit des preuves électrophysiologiques pour comprendre le mécanisme de la mémoire de travail altérée après une privation de sommeil. Il est nécessaire de prêter attention aux effets néfastes des troubles de la mémoire de travail causés par la privation de sommeil et d’explorer des interventions efficaces pour de tels dommages.
Déclaration de disponibilité des données
Les ensembles de données générés pour cette étude sont disponibles sur demande auprès de l’auteur correspondant.
Déclaration d’éthique
Les études impliquant des participants humains ont été examinées et approuvées par la Quatrième Université de médecine militaire de l’Université de Beihang. Les patients / participants ont fourni leur consentement éclairé écrit pour participer à cette étude.
Contributions des auteurs
YS a conçu les expériences. ZP a produit les résultats et a écrit le manuscrit. CD et LZ ont analysé et interprété les données. JT et YS ont effectué les expériences, ont pris connaissance des données et des garants de cette étude. YB, LZ et JT ont contribué à la collecte de données et à l’examen de la littérature. Tous les auteurs énumérés ont lu et approuvé le manuscrit final.
Financement
Cette recherche a été soutenue par la National Science Foundation of Winter Olympics Technology Plan of China dans le cadre des subventions Nos 2019YFF0301600 et HJ20191A020135.
Conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de relations commerciales ou financières pouvant être interprétées comme un conflit d’intérêts potentiel.
Anderson, C., et Platten, C. R. (2011). La privation de sommeil réduit l’inhibition et améliore l’impulsivité aux stimuli négatifs. Se comporter. Cerveau Rés. 217:466. doi: 10.1016/j.bbr.2010.09.020
Résumé PubMed |Texte intégral CrossRef/Google Scholar
Baddeley, A. (2000). Le tampon épisodique: un nouveau composant de la mémoire de travail? Tendances Cogn. Sci. 4:417. doi: 10.1016/s1364-6613(00) 01538-2
CrossRef Texte intégral/Google Scholar
Bartel, P., Offermeier, W., Smith, F. et Becker, P. (2004). Attention et mémoire de travail chez les anesthésistes résidents après le service de nuit: effets de groupe et individuels. Occupp. Environ. Med. 61, 167–170. doi: 10.1136/ oem.2002.006197
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Casement, MD, Broussard, JL, Mullington, JM, et Press, Dz (2006). La contribution du sommeil à l’amélioration de la vitesse de balayage de la mémoire de travail: une étude de la restriction prolongée du sommeil. Biol. Psychol. 72:212. doi: 10.1016 / j. biopsycho.2005.11.002
Résumé PubMed /Texte intégral CrossRef/Google Scholar
Chee, M. W. L., et Chuah, L. Y. M. (2008). La neuroimagerie fonctionnelle permet de comprendre comment le sommeil et la privation de sommeil affectent la mémoire et la cognition. Curr. Opin. Neurol. 21, 417–423. doi: 10.1097 / OMD.0b013e3283052cf7
Résumé publié/Texte intégral croisé/Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S. et Chee, M. W. L. (2005). Dissociation des régions corticales modulées à la fois par la charge de mémoire de travail et la privation de sommeil et par la privation de sommeil seule. Neuroimage 25, 579-587. doi: 10.1016/j. neuroimage.2004.11.029
Résumé PubMed / Texte intégral CrossRef |Google Scholar
Chuah, Y. M. L., Venkatraman, V., Dinges, D. F. et Chee, M. W. L. (2006). La base neurale de la variabilité interindividuelle de l’efficacité inhibitrice après la privation de sommeil. J. Neurosci. 26, 7156–7162. doi: 10.1523/jneurosci.0906-06.2006
CrossRef Texte intégral/Google Scholar
Cote, K. A., Milner, C. E., Osip, S. L., Baker, M. L. et Cuthbert, B. P. (2008). Excitation et attention physiologiques pendant une semaine de restriction continue du sommeil. Physiol. Se comporter. 95, 353–364. doi: 10.1016/ j. physbeh.2008.06.016
Résumé PubMed /Texte intégral CrossRef/Google Scholar
Cote, K. A., Mondloch, C. J., Sergeeva, V., Taylor, M. et Semplonius, T. (2014). Impact de la privation totale de sommeil sur le traitement neuronal comportemental des visages émotionnellement expressifs. Exp. Brain Res. 232, 1429-1442. doi: 10.1007/s00221-013-3780-1
Résumé PubMed | Texte intégral CrossRef /Google Scholar
Donchin, E., et Fabiani, M. (1991). L’utilisation des potentiels cérébraux liés aux événements dans l’étude de la mémoire: p300 est-il une mesure du caractère distinctif des événements? Psychol. Facul. Public. 7, 58–77. doi: 10.1111/sltb.12055
Résumé PubMed / CrossRef Texte intégral /Google Scholar
Drummond, S. P., Brown, G. G., Salamat, J. S. et Gillin, J. C. (2004). L’augmentation de la difficulté de la tâche facilite la réponse compensatoire cérébrale à la privation totale de sommeil. Dormir 27, 445-451. doi: 10.1093/sleep/27.3.445
CrossRef Texte intégral/Google Scholar
Drummond, S. P.A., et Brown, G.G. (2001). Les effets de la privation totale de sommeil sur les réponses cérébrales aux performances cognitives. Neuropsychopharmacologie 25, S68–S73. doi: 10.1016/S0893-133X(01) 00325-6
CrossRef Texte intégral/Google Scholar
Evans, K. M., et Federmeier, K.D. (2007). La mémoire qui est à droite et la mémoire qui est à gauche: les potentiels liés aux événements révèlent des asymétries hémisphériques dans le codage et la rétention des informations verbales. Neuropsychologia 45, 1777-1790. doi: 10.1016 / j. neuropsychologie.2006.12.014
Résumé publié|Texte intégral croisé/Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M. et Höller, Y. (2007). La composante p2 visuelle est liée au verrouillage de phase thêta. Neurosci. Lett. 426, 181–186. doi: 10.1016 / j.neulet.2007.08.062
Résumé PubMed | CrossRef Texte intégral/Google Scholar
Géraldine, R., Desgranges, B., Foret, J., et Eustache, F. (2005). Les relations entre les systèmes de mémoire et les stades du sommeil. J. Sommeil Rés. 14, 123-140. doi: 10.1111/j.1365-2869.2005.00450.x
Résumé PubMed | Texte intégral croisé /Google Scholar
Gosselin, A., Koninck, J. D., et Campbell, K. B. (2005). Privation totale de sommeil et traitement de la nouveauté: implications pour le fonctionnement du lobe frontal. Clin. Neurophysiol. 116, 211–222. doi: 10.1016 / j. clinph.2004.07.033
Résumé PubMed | Texte intégral CrossRef /Google Scholar
Harrison, Y. et Horne, J. (1998). La perte de sommeil nuit aux tâches langagières courtes et nouvelles ayant une focalisation préfrontale. J. Sommeil Rés. 7, 95-100. doi: 10.1046/j.1365-2869.1998.00104.x
Résumé publié/Texte intégral croisé /Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J., et Meier, B. (2010). La validité simultanée de la tâche n-back en tant que mesure de mémoire de travail. Mémoire 18, 394-412. doi: 10.1080/09658211003702171
Résumé publié|Texte intégral croisé/Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., et Chamari, K. (2013). L’impact de la privation partielle de sommeil sur les variations diurnes des performances cognitives chez les sujets entraînés. Proc. Soc. Se comporter. Sci. 82, 392–396. doi: 10.1016/j.sbspro.2013.06.281
CrossRef Texte intégral/Google Scholar
Jennings, J. R., Monk, T. H., et Molen, M. W. V. D. (2003). La privation de sommeil influence certains processus d’attention de supervision, mais pas tous. Psychol. Sci. 14, 473–479. doi: 10.1111/1467-9280.02456
Résumé publié|Texte intégral croisé /Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L. et Lei, Y. (2015). Le sommeil de récupération inverse l’inhibition de la réponse altérée due à la restriction du sommeil: preuves d’une étude des potentiels liés aux événements visuels. PLoS Un 10:e0142361. doi: 10.1371 / journal.pone.0142361
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Jones, K., et Harrison, Y. (2001). Fonction du lobe frontal, perte de sommeil et sommeil fragmenté. Dormir Med. Apoc.5:475. doi: 10.1053/smrv.2001.0203
Résumé PubMed |Texte intégral CrossRef/Google Scholar
Koslowsky, M., et Babkoff, H. (1992). Méta-analyse de la relation entre la privation totale de sommeil et la performance. Chronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
Résumé publié|Texte intégral croisé /Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A. et Hansenne, M. (2014). Abus d’alcool et composants erp dans les tâches go / no-go utilisant des stimuli liés à l’alcool: impact de l’évitement de l’alcool. Int. J. Psychophysiol. 94, 92–99. doi: 10.1016/ j. ijpsycho.2014.08.001
Résumé PubMed | Texte intégral CrossRef /Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F. et Huster, R. J. (2019). La privation de sommeil affecte différemment les sous-composantes du contrôle cognitif. Sommeil 42: zsz016. doi: 10.1093/sleep/zsz016
Résumé publié/Texte intégral croisé/Google Scholar
Lavie, P. (2001). Sommeil-réveil comme rythme biologique. Annu. Le révérend Psychol. 52, 277–303. doi: 10.1146/annurev.psy.52.1.277
CrossRef Texte intégral/Google Scholar
Lei, Y., Shao, Y., Wang, L., Zhai, T. et Zou, F. (2015). Le couplage de réseau cérébral à grande échelle prédit des effets de privation totale de sommeil sur la capacité cognitive. PLoS One 10: e0133959. doi: 10.1371 / journal.pone.0133959
Résumé publié / Texte intégral croisé /Google Scholar
Li, C., Huang, D., Qi, J., Chang, H. et Meng, Q. (2016). Les déficits de mémoire à court terme sont en corrélation avec les altérations de la connectivité fonctionnelle hippocampo-thalamique après une restriction aiguë du sommeil. L’imagerie cérébrale Se comporte. 11, 954–963. doi: 10.1007/s11682-016-9570-1
Résumé PubMed / Texte intégral CrossRef |Google Scholar
Ma, N., Dinges, D. F., Basner, M. et Rao, H. (2014). Comment la perte de sommeil totale aiguë affecte le cerveau participant: une méta-analyse des études de neuroimagerie. Dormir 38, 233. doi: 10.5665 / sommeil.4404
Résumé PubMed | Texte intégral CrossRef /Google Scholar
Miyake, A., et Shah, P. (1999). Modèles de Mémoire de Travail. Mécanismes de Maintenance Active et de Contrôle Exécutif. Cambridge : Presses universitaires de Cambridge.
Google Scholar
Mograss, M. A., Guillem, F., Brazzini-Poisson, V., et Godbout, R. (2009). Les effets de la privation totale de sommeil sur les processus de mémoire de reconnaissance: une étude du potentiel lié aux événements. Neurobiol. Apprendre. Mem. 91, 343–352. doi: 10.1016/j.nlm.2009.01.008
Résumé PubMed |Texte intégral CrossRef/Google Scholar
Montplaisir, J. (1981). Dépression et rythme biologique: effets thérapeutiques de la privation de sommeil. L’unión Médicale du Canada 110, 272-276. doi: 10.1620/ tjem.133.481
Résumé PubMed /CrossRef Texte intégral/Google Scholar
Morris, A. M., So, Y., Lee, K. A., Lash, A. A., et Becker, C.E. (1992). Le potentiel lié à l’événement p300. les effets de la privation de sommeil. J. Occupat. Med. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F., et Hobson, J.A. (2002). Le cortex préfrontal dans le sommeil. Tendances Cogn. Sci. 6, 475–481. doi: 10.1016/S1364-6613(02) 01992-7
CrossRef Texte intégral/Google Scholar
Ning, M., Dinges, D. F., Mathias, B. et Hengyi, R. (2014). Comment la perte de sommeil totale aiguë affecte le cerveau participant: une méta-analyse des études de neuroimagerie. Dormir 38, 233-240. doi: 10.5665 / sommeil.4404
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, A. R. et Bullmore, E. (2005). Paradigme de la mémoire de travail N-back: une méta-analyse des études de neuroimagerie fonctionnelle normative. Hum. Mapp cerveau. 25, 46–59. doi: 10.1002/hbm.20131
Résumé PubMed | Texte intégral CrossRef /Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S. et Kumar, S. (2010). Les corrélats électrophysiologiques de la cognition s’améliorent avec la sieste pendant la privation de sommeil. EUR. J. Appl. Physiol. 108, 549–556. doi: 10.1007/s00421-009-1222-3
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Ratcliff, R., et Van Dongen, H. (2018). Les effets de la privation de sommeil sur l’élément et la mémoire de reconnaissance associative. J. Exp. Psychol. 44, 193–208. doi: 10.1037/xlm0000452
Résumé publié/Texte intégral croisé/Google Scholar
Raymond, C.A. (1988). Le changement de travail, les cycles de sommeil sont en passe de devenir un autre problème de santé publique. JAMA J. Am. Med. Assoc. 259, 2958–2959. doi: 10.1001/ jama.259.20.2958
CrossRef Texte intégral/Google Scholar
Saito, H., Yamazaki, H., Matsuoka, H., Matsumoto, K. et Sato, M. (2001). Potentiel lié aux événements visuels dans la démence légère de type Alzheimer. Clin de psychiatrie. Neurosci. 55, 365–371. doi: 10.1046 / j.1440-1819.2001.00876.x
Résumé publié/Texte intégral croisé/Google Scholar
Schacht, A., Werheid, K., et Sommer, W. (2008). L’évaluation de la beauté du visage est rapide mais pas obligatoire. Cogn. Affecter. Se comporter. Neurosci. 8, 132–142. doi: 10.3758/ cabn.8.2.132
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Scott, J. P. R., Mcnaughton, L. R., et Polman, R. C. J. (2006). Effets de la privation de sommeil et de l’exercice sur les performances cognitives, motrices et l’humeur. Physiol. Se comporter. 87:408. doi: 10.1016/ j. physbeh.2005.11.009
Résumé PubMed /CrossRef Texte intégral/Google Scholar
Smallwood, J., Brown, K., Baird, B., et Schooler, J. W. (2011). Coopération entre le réseau de mode par défaut et le réseau fronto-pariétal dans la production d’un train de pensée interne. Brain Res. 1428, 60-70. doi: 10.1016 / j. brainres.2011.03.072
Résumé PubMed /Texte intégral CrossRef/Google Scholar
Smith, M. E., Mcevoy, L. K. et Gevins, A. (2002). L’impact d’une perte de sommeil modérée sur les signaux neurophysiologiques pendant l’exécution des tâches de mémoire de travail. Dormir 25, 784-794. doi: 10.1093 / sommeil / 25.7.56
CrossRef Texte intégral/Google Scholar
Thomas, M. (2003). Base neuronale de la vigilance et des troubles de la performance cognitive pendant la somnolence ii. effets de 48 et 72 h de privation de sommeil sur l’activité cérébrale régionale humaine éveillée. Thalam. Relat. Syst. 2, 199–229. doi: 10.1016/S1472-9288(03) 00020-7
CrossRef Texte intégral/Google Scholar
Van Dongen, H. (2004). Le coût cumulé de l’éveil supplémentaire: effets de la réponse doseuse sur les fonctions neurocomportementales et la physiologie du sommeil de la privation chronique de sommeil et de la privation totale de sommeil. Sommeil 26, 117-126. doi: 10.1016/0739-6260(84)90040-6 il s’agit d’un outil de recherche et d’analyse de données qui permet d’identifier et d’analyser les données de l’utilisateur. La privation de sommeil entraîne une perte de connectivité fonctionnelle dans les régions frontales du cerveau. BMC Neurosci. 15:88. doi: 10.1186|1471-2202-15-88
Résumé publié/Texte intégral croisé/Google Scholar
Wen, X., Liu, Y., Yao, L., et Ding, M. (2013). Régulation descendante de l’activité du mode par défaut dans l’attention visuelle spatiale. J. Neurosci. 33, 6444–6453. doi: 10.1523/JNEUROSCI.4939-12.2013
CrossRef Texte intégral/Google Scholar
Wiggins, E., Malayka, M. et Kendra, G. (2018). la privation de sommeil de 24 heures altère la modulation de l’attention précoce du traitement neuronal: une étude du potentiel cérébral liée aux événements. Neurosci. Lett. 677, 32–36. doi: 10.1016 / j.neulet.2018.04.022
Résumé publié / Texte intégral croisé /Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y. et Zhou, Q. (2019). Diminution du remplacement de l’information de la mémoire de travail après une privation de sommeil: preuves d’une étude potentielle liée à un événement. Devant. Neurosci. 13:408. doi: 10.3389/ fnins.2019.00408
CrossRef Texte intégral /Google Scholar
Zhang, Q., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z., et al. (2014). Une étude erp visuelle de l’inhibition des impulsions suite à une sieste induite par le zaleplon après une privation de sommeil. PLoS One 9: e95653. doi: 10.1371 / journal.pone.0095653
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Leave a Reply