Bibliothèque
Malgré sa localisation périphérique, la rétine ou partie neurale de l’œil, fait en fait partie du système nerveux central. Au cours du développement, la rétine se forme comme une sortie du diencéphale, appelée vésicule optique, qui subit une invagination pour former la cupule optique (Figure 11.3; voir aussi Chapitre 22). La paroi interne de la cupule optique donne naissance à la rétine, tandis que la paroi externe donne naissance à l’épithélium pigmentaire. Cet épithélium est une structure contenant de la mélanine qui réduit la rétrodiffusion de la lumière qui pénètre dans l’œil; il joue également un rôle essentiel dans le maintien des photorécepteurs, le renouvellement des photopigments et la phagocytose des disques photorécepteurs, dont le renouvellement à un taux élevé est essentiel à la vision.

Figure 11.3
Développement de l’œil humain. (A) La rétine se développe comme une sortie du tube neural, appelée vésicule optique. (B) La vésicule optique s’invagine pour former la coupelle optique. (C, D) La paroi interne de la cupule optique devient la rétine neurale, tandis que le (more…)
Conformément à son statut de partie à part entière du système nerveux central, la rétine comprend des circuits neuronaux complexes qui convertissent l’activité électrique graduée des photorécepteurs en potentiels d’action qui se déplacent vers le cerveau via les axones du nerf optique. Bien qu’elle ait les mêmes types d’éléments fonctionnels et de neurotransmetteurs que dans d’autres parties du système nerveux central, la rétine ne comprend que quelques classes de neurones, et ceux-ci sont disposés de manière moins difficile à démêler que les circuits dans d’autres zones du cerveau. Il existe cinq types de neurones dans la rétine: les photorécepteurs, les cellules bipolaires, les cellules ganglionnaires, les cellules horizontales et les cellules amacrines. Les corps cellulaires et les processus de ces neurones sont empilés en cinq couches alternées, les corps cellulaires étant situés dans les couches cellulaires nucléaire interne, nucléaire externe et ganglionnaire, et les processus et les contacts synaptiques situés dans les couches plexiforme interne et plexiforme externe (Figure 11.4). Une chaîne directe à trois neurones – cellule photoréceptrice à cellule bipolaire à cellule ganglionnaire – est la principale voie de circulation de l’information des photorécepteurs vers le nerf optique.
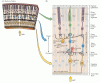
Figure 11.4
Structure de la rétine. (A) Section de la rétine montrant la disposition générale des couches rétiniennes. (B) Schéma des circuits de base de la rétine. Une chaîne à trois neurones – photorécepteur, cellule bipolaire et cellule ganglionnaire – fournit le plus direct (plus…)
Il existe deux types d’éléments sensibles à la lumière dans la rétine: les bâtonnets et les cônes. Les deux types de photorécepteurs ont un segment externe composé de disques membraneux contenant un photopigment et adjacents à la couche épithéliale pigmentaire, et un segment interne contenant le noyau cellulaire et donnant naissance à des terminaux synaptiques qui entrent en contact avec des cellules bipolaires ou horizontales. L’absorption de la lumière par le photopigment dans le segment externe des photorécepteurs déclenche une cascade d’événements qui modifie le potentiel membranaire du récepteur, et donc la quantité de neurotransmetteur libérée par le photorécepteur se synapse sur les cellules qu’ils entrent en contact. Les synapses entre les terminaux photorécepteurs et les cellules bipolaires (et les cellules horizontales) se produisent dans la couche plexiforme externe; plus précisément, les corps cellulaires des photorécepteurs constituent la couche nucléaire externe, tandis que les corps cellulaires des cellules bipolaires se trouvent dans la couche nucléaire interne. Les processus axonaux courts des cellules bipolaires établissent des contacts synaptiques à leur tour sur les processus dendritiques des cellules ganglionnaires de la couche plexiforme interne. Les axones beaucoup plus grands des cellules ganglionnaires forment le nerf optique et transmettent des informations sur la stimulation rétinienne au reste du système nerveux central.
Les deux autres types de neurones de la rétine, les cellules horizontales et les cellules amacrines, ont leurs corps cellulaires dans la couche nucléaire interne et sont principalement responsables des interactions latérales au sein de la rétine. Ces interactions latérales entre les récepteurs, les cellules horizontales et les cellules bipolaires de la couche plexiforme externe sont en grande partie responsables de la sensibilité du système visuel au contraste de luminance sur une large gamme d’intensités lumineuses. Les processus des cellules amacrines, qui s’étendent latéralement dans la couche plexiforme interne, sont postsynaptiques aux terminaisons cellulaires bipolaires et présynaptiques aux dendrites des cellules ganglionnaires (voir Figure 11.4). Les processus des cellules horizontales se ramifient dans la couche plexiforme externe. Plusieurs sous-classes de cellules amacrines qui apportent des contributions distinctes à la fonction visuelle. Une classe de cellules amacrines, par exemple, joue un rôle important dans la transformation des réponses persistantes des cellules bipolaires à la lumière en réponses transitoires brèves exposées par certains types de cellules ganglionnaires. Un autre type sert d’étape obligatoire dans la voie qui transmet les informations des photorécepteurs de bâtonnets aux cellules ganglionnaires rétiniennes. La variété des sous-types de cellules amacrines illustre la règle plus générale selon laquelle, bien qu’il n’existe que cinq types de cellules rétiniennes de base, il peut y avoir une diversité considérable au sein d’un type de cellules donné. Cette diversité est à la base des voies qui transmettent différentes sortes d’informations aux cibles centrales de manière parallèle.
À première vue, la disposition spatiale des couches rétiniennes semble contre-intuitive, car les rayons lumineux doivent traverser les éléments non sensibles à la lumière de la rétine (et du système vasculaire rétinien!) avant d’atteindre les segments externes des photorécepteurs, où les photons sont absorbés (voir Figure 11.4). La raison de cette curieuse caractéristique de l’organisation rétinienne réside dans la relation particulière qui existe entre les segments externes des photorécepteurs et l’épithélium pigmentaire. Les segments externes contiennent des disques membraneux qui abritent le photopigment sensible à la lumière et d’autres protéines impliquées dans le processus de transduction. Ces disques sont formés près du segment interne du photorécepteur et se déplacent vers l’extrémité du segment externe, où ils sont perdus. L’épithélium pigmentaire joue un rôle essentiel dans l’élimination des disques récepteurs dépensés; ce n’est pas une mince tâche, car tous les disques des segments externes sont remplacés tous les 12 jours. De plus, l’épithélium pigmentaire contient la machinerie biochimique nécessaire à la régénération des molécules de photopigment après leur exposition à la lumière. Ce sont vraisemblablement les exigences du cycle de vie du disque photorécepteur et du recyclage des photopigments qui expliquent pourquoi les bâtonnets et les cônes se trouvent dans la couche la plus externe plutôt que la plus interne de la rétine. Les perturbations des relations normales entre l’épithélium pigmentaire et les photorécepteurs rétiniens, telles que celles qui surviennent dans la rétinite pigmentaire, ont de graves conséquences sur la vision (encadré B).
Box B
Retinitis Pigmentosa.
Leave a Reply