Fronteras en Neurociencia
Introducción
Con el progreso de la sociedad y los cambios en el ritmo de trabajo, un número creciente de personas sufre de privación de sueño. La privación del sueño no solo daña la salud física y mental del individuo, sino que también afecta gravemente el rendimiento laboral, causando errores de trabajo e incluso accidentes. Por lo tanto, comprender el mecanismo de privación del sueño que afecta la función cognitiva es de gran importancia para prevenir eficazmente los efectos de la privación del sueño.
Estudios previos han revelado que la privación del sueño puede causar una serie de cambios en el estado de ánimo, la capacidad cognitiva, el rendimiento laboral y la función inmunológica de un individuo (Choo et al., 2005). La falta de sueño altera la circulación corporal y afecta las capacidades cognitivas y emocionales de los individuos (Raymond, 1988). Varios estudios han revelado que la privación del sueño afecta la inhibición de la respuesta (Harrison y Horne, 1998; Muzur et al., 2002; Jennings et al., 2003). Por ejemplo, después de 36 h de privación de sueño, la capacidad del individuo para suprimir los estímulos negativos disminuyó (Chuah et al., 2006). Los estudios de neuroimagen han sugerido que la privación del sueño reduce el bajo nivel de capacidad de procesamiento visual de un individuo (Anderson y Platten, 2011; Ning et al., 2014). Además, la privación del sueño afecta el hipocampo y podría afectar la memoria al destruir la plasticidad sináptica(Cote et al., 2014). Thomas (2003) ha indicado que la falta de sueño reduce el flujo sanguíneo cerebral y la tasa metabólica en el tálamo, la corteza prefrontal y la corteza parietal (Géraldine et al., 2005). Jarraya y sus colegas descubrieron que la privación parcial del sueño afectaba significativamente las funciones neuropsicológicas, como la memoria verbal instantánea, la atención y el estado de alerta (Thomas, 2003). Además, algunos estudios han revelado que los efectos acumulativos de la privación parcial del sueño podrían perjudicar gravemente la función cognitiva y el comportamiento (Van Dongen, 2004; Scott et al., 2006; Jarraya et al., 2013).
La memoria de trabajo es un sistema que se utiliza para almacenar y procesar información y que es una función cognitiva con capacidad limitada (Bartel et al., 2004). Además, la información almacenada en el sistema de memoria de trabajo se puede cambiar de memoria a corto plazo a memoria a largo plazo a través de recuentos y otros métodos de memoria. La memoria de trabajo es la transición entre sistemas de memoria a corto y largo plazo, que es muy fundamental en el procesamiento de mensajes humanos (Miyake y Shah, 1999). Proporciona un espacio de almacenamiento temporal y los recursos necesarios para procesar la información, como la comprensión de la voz, el razonamiento y el aprendizaje. Se ha demostrado que la privación del sueño afecta primero a la memoria de trabajo.
Estudios anteriores han utilizado el paradigma de la memoria de trabajo n-back en participantes que se sometieron a privación de sueño y encontraron que la falta de sueño induce una disminución de la actividad metabólica en la red regional del cerebro, que se efectúa principalmente procesamiento de información e inhibición de reacciones (Baddeley, 2000; Zhang et al., 2019). El deterioro de la memoria de trabajo después de la privación del sueño está relacionado con la activación de la red predeterminada en las tareas (Chee y Chuah, 2008), lo que puede estar relacionado con el importante papel del tálamo en el estado de alerta cortical. Por ejemplo, la privación de sueño aumentó la conexión entre el hipocampo, el tálamo y la red predeterminada, lo que a menudo iba acompañado de una mayor somnolencia subjetiva y un peor rendimiento de la memoria de trabajo (Lei et al., 2015; Li et al., 2016). ¿Estudios sobre la identidad de la privación de sueño?de que el aumento de la latencia y la amplitud reducida del componente P3 se asociaron con una sobriedad prolongada (Morris et al., 1992; Jones y Harrison, 2001; Panjwani et al., 2010). La disminución en la onda P3 podría reflejar una disminución en la atención de los participantes y una reducción en el discernimiento de los estímulos objetivo (Koslowsky y Babkoff, 1992).
Sin embargo, pocos estudios han proporcionado evidencia electrofisiológica de deterioro de la memoria de trabajo después de la privación del sueño. La tarea n-back se considera un método común para evaluar la memoria de trabajo (Owen et al., 2005; Jaeggi et al., 2010). Zhang et al. diseñó una tarea de memoria de trabajo de pronunciación de dos columnas para explorar la alternativa de mensajes disminuidos de la memoria de trabajo durante la privación de sueño, pero pocos estudios han utilizado diferentes tipos de tareas de memoria de trabajo en un solo experimento. En el presente estudio, diseñamos diferentes tipos de tareas de memoria de trabajo (memoria de trabajo de pronunciación, memoria de trabajo espacial y memoria de trabajo de objetos) para explorar el deterioro de la función cognitiva por TSD y los datos registrados del EEG de los participantes en 2 puntos de tiempo (línea de base y 36 h-TSD). Todas las tareas adoptaron un paradigma de dos partes. Este estudio evaluó los cambios en la onda N2-P3 relacionados con la memoria de trabajo durante la TSD y analizó las características temporales de los efectos de la privación del sueño en la memoria de trabajo. Nuestros hallazgos proporcionan evidencia experimental de los efectos de la privación del sueño en la función cognitiva.
Materiales y métodos
Participantes
Dieciséis estudiantes varones jóvenes, sanos y diestros participaron en este estudio. Reclutamos participantes mediante publicidad en el campus. Todos los participantes tenían buenos hábitos de sueño (PSQI< 5). Todos los participantes tenían entre 21 y 28 años con una edad promedio de 23 años, y ninguno de los participantes tenía ninguna enfermedad mental o física. Todos los participantes tenían visión normal o visión corregida por encima de 1,0 y puntuaciones de inteligencia >110 en la prueba de Raven. Antes del experimento, el experimentador explicó el procedimiento y señaló a la atención de los participantes para asegurarse de que estuvieran familiarizados con el método y el procedimiento. En las 2 semanas anteriores al experimento, los participantes dormían regularmente durante 7-9 horas al día, sin fumar, beber café, beber alcohol o consumir cualquier medicamento durante 2 días antes del experimento. Antes del experimento, todos los participantes dieron su consentimiento informado por escrito. El plan experimental fue aprobado por el Comité de Ética de la Cuarta Universidad Médica Militar y la Universidad de Beihang.
Diseño experimental
Se presentaron a todos los participantes tres tipos de tareas de memoria de trabajo. Eran dos tareas de memoria de trabajo de pronunciación (ver Figura 1), dos tareas de memoria de trabajo espacial (ver Figura 2) y dos tareas de memoria de trabajo de objeto (ver Figura 3). Los materiales de estimulación de las tareas fueron 15 letras inglesas insensibles a mayúsculas y minúsculas, excluyendo las que tienen letras similares, como L/l, M / m, cuadrados negros pequeños y 12 figuras geométricas, respectivamente. Todos los materiales se mostraron en color negro sobre un fondo blanco, con un ángulo visual aproximado de 1,5° × 1,5° (ancho: 2,0 cm, alto: 2,0 cm). se incluyeron 122 senderos en cada tarea y, en cada sendero, se presentó el estímulo objetivo para 400 ms dos senderos después de la presentación del estímulo objetivo, con el tiempo de asincronía de inicio del estímulo (SOA) de 1,600 ms que estaba marcado por un blanco»+».»Se pidió a los participantes que hicieran clic con el botón izquierdo del ratón cuando el estímulo objetivo y el objetivo fueran iguales («coincidentes»), mientras que hicieran clic con el botón derecho del ratón cuando no lo fueran («no coincidentes»). La condición de coincidencia o no se presentó en un orden pseudoaleatorio con una proporción de 1: 1.
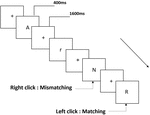
Figura 1. Diagrama esquemático de la tarea de memoria de trabajo de pronunciación.
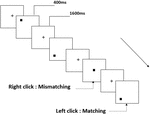
la Figura 2. Diagrama esquemático de la tarea de memoria de trabajo espacial.
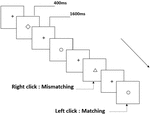
la Figura 3. Diagrama esquemático de la tarea de memoria de trabajo del objeto.
Procedimientos experimentales
Antes del experimento, se instruyó a los participantes de la tarea experimental. Se les informó que practicaran los tres tipos de tareas de memoria de trabajo hasta que se lograra una tasa de precisión del 90%. Los participantes visitaron el laboratorio el día anterior al experimento y durmieron en el laboratorio esa noche. Los dos socios participantes realizaron los experimentos al mismo tiempo. Se realizaron tres tipos de tareas de memoria de trabajo de 7:30 a.m. a 8:30 a. m. a la mañana siguiente con registro simultáneo de electroencefalograma (EEG) (línea de base). La segunda grabación de EEG (36 h-TSD) se realizó después de un período de 36 h durante el cual los participantes no pudieron dormir. Durante todo el tiempo del experimento, se prohibieron la inhibición central y los estimulantes. Los participantes fueron acompañados, observados y recordados por el personal de enfermería para mantenerlos despiertos durante toda la sesión de TSD.
Grabaciones de EEG
Se grabó un EEG continuo del cuero cabelludo utilizando tapas de electrodos colocadas en 64 ubicaciones utilizando el sistema 10-20 con un amplificador SynAmps2. Las mastoides bilaterales (A1 y A2) se usaron como referencia, y la frente se usó como suelo. Los EEG se registraron a 1.000 Hz y la impedancia de todos los canales se mantuvo por debajo de 5 kΩ. Se colocaron cuatro electrodos adicionales por encima y por debajo de los ojos derecho e izquierdo para registrar un electrooculograma bipolar vertical y horizontal.
Análisis de datos de Experimentos Conductuales
Debido a errores técnicos, se eliminaron dos casos y se incluyeron otros 14 casos en el siguiente análisis estadístico. Los datos de comportamiento incluyeron el tiempo de reacción medio, la tasa correcta y el número correcto por unidad de tiempo. Se registraron datos de comportamiento en los estados basales y 36 h-TSD para su análisis. Los análisis fueron realizados por IBM SPSS (V22.2), donde se lanzó el método de análisis de varianza de medidas repetidas (ANOVA) con Geisser de Efecto Invernadero fue el análisis post hoc Bonferroni. Los resultados estadísticos se presentaron como media y desviación estándar (DE).
Análisis de datos de EEG
Se utilizó el programa Scan 4.3 para analizar los datos de EEG, donde los artefactos de EEG del movimiento ocular se corrigieron mediante el método de reducción de artefactos oculares. Las épocas que oscilaban entre -100 y 800 ms de los datos continuos del EEG se extrajeron y filtraron mediante un filtro de paso de banda de 0,5 a 30 Hz con una pendiente de frecuencia de 24 dB/oct. Se rechazaron los ensayos en los que la tensión excedió ± 100 µV y se corrigió el valor basal a una amplitud media de 100 ms. Los componentes del EEG se promediaron y calcularon solo con las respuestas corregidas. Se identificaron y cuantificaron los componentes de ERP P2 (100-250 ms), N2 (150-350 ms) y P3 (250-450 ms) de los ensayos de estímulo. Las amplitudes y latencias de pico de gran promedio de los componentes N2 y P3 se calcularon por separado en F3, Fz, F4, C3, Cz, C4, P3, Pz y P4, y el componente P2 se calculó en F3, Fz, F4, C3, Cz y C4 (Casement et al., 2006; Verweij et al., 2014).
Se emplearon medidas repetidas ANOVA para todos los resultados de ERP. Los principales efectos y las interacciones entre estados de sueño (línea de base y 36 h-TSD), tareas (memoria de trabajo de pronunciación, memoria de trabajo espacial y memoria de trabajo de objetos), regiones (frontal, central y parietal; el componente P2 se analizó solo en las regiones frontal y central) y sitios (izquierda, central y derecha) se analizaron estadísticamente utilizando medidas repetidas ANOVA, que incluyeron correcciones de Geisser de invernadero para pruebas post hoc de no esferroni y Bonferroni.
Resultados
Rendimiento Conductual
Los resultados de los experimentos conductuales se muestran en la Tabla 1. El tiempo medio de reacción fue mayor en el estado de 36 h-TSD que en el estado basal, con una tendencia a aumentar pero sin diferencias significativas. La ANOVA reveló que la velocidad correcta de la tarea fue significativamente diferente entre el valor basal y 36 h-TSD . El número correcto por unidad de tiempo mostró un efecto principal significativo del tiempo durante 36 h-TSD .

Cuadro 1. Datos de rendimiento (media ± DE) en la tarea de 2 espaldas al inicio y después de 36 h-TSD.
Amplitud
En comparación con el valor basal, se observó una disminución significativa de la amplitud de P3 y un aumento significativo de la amplitud de P2 después de la TSD. Aunque la amplitud de N2 disminuyó después de 36 h de TSD, la diferencia no alcanzó significación estadística (Tabla 2).

la Tabla 2. Amplitud máxima de gran promedio de los componentes P2, N2 y P3 en la condición de respuesta correcta en múltiples sitios de electrodos al inicio y después de 36 h-TSD.
Se encontraron efectos principales significativos de regiones y sitios en la amplitud P2 bajo la condición de TSD. Durante la TSD, la amplitud máxima de P2 apareció en la región frontal (Figura 4). Además, las diferencias en amplitudes de P2 en diferentes regiones (frontal vs.central) fueron más significativas en los electrodos bilaterales (izquierda: P = 0,001; derecha: P = 0,000) que en los electrodos medios (Figura 4). Un efecto principal significativo de la región y el sitio en N2 reveló que la amplitud de N2 era más negativa en el frontal que en la región central (P = 0,008, Figura 4) y era más pequeña en el lado derecho que en el lado izquierdo (P = 0,011, Figuras 5A2,B2). Se observó un efecto principal del sitio en la amplitud P3 . La amplitud de P3 fue más positiva en el medio que en el lado izquierdo (P = 0,009, Figura 4). Se observó un efecto de interacción significativo entre el tiempo y la región para la amplitud P3 . Durante la TSD, la reducción de la amplitud de P3 fue más significativa en las regiones frontal y central que en la región parietal (P = 0,005; P = 0,003) (Figura 5C3). Ningún otro efecto principal o efecto de interacción alcanzó significación estadística.
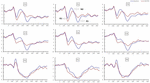
la Figura 4. Amplitud ERP en la línea de base y 36 h-TSD para la condición de respuesta correcta para la tarea de memoria de trabajo. Los canales se ordenan de izquierda a derecha y de arriba a abajo de la siguiente manera: F3, Fz, F4, C3, Cz, C4, P3, Pz y P4. En comparación con el valor basal, las latencias de los componentes N2-P3 se prolongaron y las amplitudes de N2-P3 disminuyeron después de 36 h-TSD.
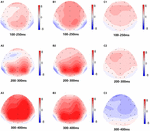
la Figura 5. Mapa topográfico de la respuesta correcta en la tarea de memoria de trabajo en diferentes condiciones de sueño (A1–C3). A1) P2, 100-250 ms, al inicio del estudio. A2) N2, 200-300 ms, al inicio del estudio. A3) P3, 300-400 ms, al inicio del estudio. B1) P2, 100-250 ms, a 36 h-TSD. B2) N2, 200-300 ms, para 36 h-TSD. B3) P3, 300-400 ms, para 36 h-TSD. (C1) P2, 100-250 ms, 36 h-TSD con el valor basal restado. (C2) N2, 200-300 ms, 36 h-TSD con el valor basal restado. (C3) P3, 300-400 ms, 36 h-TSD con el valor basal restado.
Latencia
Las latencias de N2 y P2 se prolongaron significativamente después de la TSD. Aunque la latencia de P3 se prolongó después de 36 h de TSD, la diferencia no alcanzó significación estadística (Tabla 3).

la Tabla 3. Latencia máxima de gran promedio de los componentes P2, N2 y P3 en la condición de respuesta correcta en múltiples sitios de electrodos al inicio y después de 36 h-TSD.
El efecto principal significativo de la región en N2 y P3 reveló que la latencia de los componentes N2-P3 era más corta en la región parietal que en la región frontal (P = 0,002; P = 0,000) y en la región central (P = 0,000; P = 0,000) (Figura 4). La latencia de la onda P3 fue significativamente mayor en el lado izquierdo que en el derecho (Figura 4).
Ningún otro efecto principal o efecto de interacción alcanzó significación estadística.
Las amplitudes y latencias N2, P2 y P3 que se obtuvieron en los nueve sitios de electrodos se presentan en la Figura 4. El mapa topográfico de la respuesta correcta en la tarea de memoria de trabajo en diferentes condiciones de sueño (basal, 36 h-TSD y la diferencia entre las dos condiciones) se presenta en la Figura 5.
Discusión
En este estudio, reportamos las influencias de la privación de sueño de 36 h en la memoria de trabajo, combinando datos de comportamiento en dos estados de sueño (basal y 36 h-TSD) con registros de EEG contemporáneos. El análisis de los resultados indicó que los cambios en los datos de comportamiento de acuerdo con la memoria de trabajo deteriorada después de 36 h de TSD: un aumento en el tiempo de reacción promedio de las tareas cognitivas y una disminución en la precisión.
La privación del sueño afectó el control individual de los recursos de atención. Aunque los individuos trataron de mantener la vigilia y el rendimiento en el trabajo, incluido el tiempo de reacción y la velocidad correcta, durante la privación del sueño, la capacidad de procesamiento de la información de su memoria de trabajo todavía se vio afectada debido a la disminución en la velocidad de procesamiento de la información (Casement et al., 2006; Wiggins et al., 2018). En este estudio, se midieron las ondas N2 y P3 relacionadas con la memoria de trabajo para mostrar un aumento de la latencia y una disminución de la amplitud después de la privación del sueño en comparación con las lecturas basales. Los estudios han demostrado que la privación del sueño conduce a una disminución continua de la atención, y el fenómeno de la disminución de la amplitud P3 indica que el control de la cognición de arriba hacia abajo de los individuos colapsa gradualmente. La privación del sueño tiene un efecto más adverso en las funciones cognitivas, especialmente en aquellas que dependen de las cogniciones o mentales (Kusztor et al., 2019).
El componente P3 refleja el despliegue de recursos de atención, y la latencia de P3 se ve ampliamente como la ventana de tiempo para la categorización y evaluación de estímulos. La disminución en la amplitud de la onda P3 también confirmó que la toma de decisiones en la respuesta de coincidencia después de la TSD había sido dañada en cierta medida (Gosselin et al., 2005). Los estudios han sugerido que la privación de sueño puede afectar la etapa de procesamiento de información de la memoria de trabajo. En este estudio, los indicadores de desempeño también respaldaron la conclusión de que el tiempo de respuesta al estímulo objetivo se incrementó y que la latencia de la onda P3 se prolongó (Cote et al., 2008). Se especuló que el efecto de la privación de sueño en los componentes P3 también podría tener lugar debido a la falta de respuesta al cambio de información, lo que es consistente con las conclusiones anteriores de que los componentes P3 están relacionados con la actualización del contenido de la memoria de trabajo (Donchin y Fabiani, 1991).
Estudios previos han considerado el componente N2 como un índice electrofisiológico que refleja la capacidad del individuo para suprimir la respuesta (Kreusch et al., 2014). Después de la privación del sueño, la latencia prolongada del componente NoGo-N2 indica que la capacidad del individuo para suprimir la respuesta se ve afectada (Jin et al., 2015). La disminución de la amplitud y la latencia prolongada del componente N2 relacionado con la memoria de trabajo de pronunciación después de la privación de sueño revela que la privación de sueño afecta el procesamiento de la información de la memoria de trabajo de pronunciación (Zhang et al., 2019). Generalmente se cree que el componente N2 refleja la atención selectiva del cerebro y el procesamiento de estímulos o señales emocionales (Schacht et al., 2008) y es un componente endógeno relacionado con el estado mental, la atención y el grado de atención de un individuo. En este estudio, encontramos que la latencia del componente N2 se prolongó significativamente, pero la amplitud mostró solo una tendencia a la baja. Según estudios anteriores, la latencia prolongada de N2 reflejaba un aumento del tiempo de respuesta después de la restricción del sueño (Zhang et al., 2014). Sin embargo, el hallazgo de que la amplitud de N2 no se alteró significativamente puede deberse a respuestas compensatorias cerebrales (Drummond y Brown, 2001). En el caso de recursos cognitivos limitados, había un mecanismo de compensación para restaurar la función cognitiva deteriorada(Jin et al., 2015).
De acuerdo con la topografía del cuero cabelludo, los cambios en los componentes N2-P3 relacionados con la privación del sueño son más evidentes en el área frontal. El control frontoparietal (CPF) juega un papel importante en el control cognitivo. Los estudios han demostrado que el CPF puede eludir el control cognitivo de arriba hacia abajo, lo que permite a los individuos centrarse en la información relacionada con el objetivo mientras suprime la información que no está relacionada con el objetivo (Smallwood et al., 2011; Wen et al., 2013). El CPF es importante para la retención y el procesamiento de la información en la memoria de trabajo, y el grado de activación del CPF después de la privación del sueño se redujo en comparación con un grupo de sueño normal (Ma et al., 2014). Aunque los resultados del EEG no reflejaron en detalle los cambios en regiones cerebrales específicas, reflejaron intuitivamente el efecto de la TSD en la retención y el procesamiento de la información de la memoria de trabajo.
Aunque el proceso cognitivo exacto en el que se basa el componente P2 todavía es ampliamente debatido, como una definición amplia, el componente P2 refleja el proceso de atención y procesamiento visual y generalmente se considera relacionado con la atención selectiva y la memoria de trabajo, lo que refleja el juicio temprano del proceso perceptivo (Saito et al., 2001). En este estudio, encontramos un aumento significativo en la amplitud de la onda P2 después de la privación del sueño. Los estudios han informado que las ondas P2 que podrían ser parte del sistema de emparejamiento cognitivo temprano para el procesamiento de mensajes y pueden comparar las entradas sensoriales con la memoria almacenada (Freunberger et al., 2007) son sensibles a las alteraciones en la atención de la misión y las demandas de memoria de trabajo (Smith et al., 2002). La compensación funcional es una de las funciones únicas del cerebro humano y un factor importante para mantener la función cognitiva. La activación excesiva de la corteza prefrontal dorsolateral (DLPFC) después de la privación del sueño indica que, a medida que disminuyen los recursos cerebrales, la DLPFC parece tener una función compensatoria (Drummond et al., 2004; Choo et al., 2005). Por lo tanto, se especula que el aumento significativo de la amplitud de P2 observado en este estudio puede deberse a una compensación funcional en la que los individuos parecen mantener la función cognitiva normal después de la privación del sueño. Aunque un gran número de estudios han utilizado la tecnología ERP para explorar el efecto de la privación de sueño en las funciones cognitivas, los componentes tempranos como N1 y P2 no se han estudiado sistemáticamente, y los resultados son inconsistentes (Evans y Federmeier, 2007; Wiggins et al., 2018; Zhang et al., 2019). Hay pocas investigaciones que exploren el cambio del componente P2 durante la privación del sueño(Mograss et al., 2009). Por lo tanto, los efectos de la privación de sueño en los componentes tempranos de ERP, como P2, todavía necesitan ser estudiados y explorados más a fondo.
En este experimento, utilizamos el modelo de 2 columnas para diseñar tareas de memoria de trabajo de pronunciación, espacial y de objetos y examinamos el deterioro de la memoria de trabajo después de 36 h de TSD. En comparación con estudios anteriores que se centraron solo en los efectos de la privación del sueño en un tipo específico de información, como la pronunciación, la memoria de trabajo o la función cognitiva específica, como la inhibición de la respuesta, hemos considerado el contenido del modelo de memoria de trabajo y analizado exhaustivamente los efectos de la privación del sueño en la memoria de trabajo.
sin Embargo, el estudio tiene algunas limitaciones. En primer lugar, solo usamos la tarea de 2 vueltas y no pudimos comparar el rendimiento de los participantes en tareas de memoria de trabajo de diferentes dificultades. Por lo tanto, existen limitaciones a la hora de explicar e inferir los cambios en el volumen de trabajo. En segundo lugar, solo se utilizaron voluntarios varones en el estudio, y las conclusiones deben evaluarse al extenderlas a las voluntarias. Debido al número limitado de participantes, solo encontramos que la amplitud de la onda N2 tenía una tendencia descendente y que la latencia de la onda P3 tenía una tendencia prolongada. Se pueden obtener resultados estables después de aumentar el número de participantes. En tercer lugar, la combinación de nuestro procedimiento con fMRI para la memoria de trabajo puede facilitar la interpretación de los resultados. Estudios previos han demostrado que los biorritmos circadianos afectan el desempeño conductual, y hay diferencias individuales (Montplaisir, 1981; Lavie, 2001). No registramos los datos del EEG al mismo tiempo en este experimento, por lo que no se puede descartar por completo la influencia de los biorritmos circadianos en los resultados de las pruebas.
Esta investigación mostró que la capacidad de memoria de trabajo se ve afectada después de la TSD y que este daño no se asocia con el contenido de estímulo de la memoria de trabajo. La falta de sueño redujo la calidad de la información almacenada en la memoria, lo que podría ocurrir con el proceso degenerativo de la atención (Ratcliff y Van Dongen, 2018). Este estudio proporciona evidencia electrofisiológica para comprender el mecanismo bajo la memoria de trabajo deteriorada después de la privación del sueño. Es necesario prestar atención a los efectos adversos del deterioro de la memoria de trabajo causado por la falta de sueño y explorar intervenciones eficaces para tales daños.
Declaración de disponibilidad de datos
Los conjuntos de datos generados para este estudio están disponibles a petición del autor correspondiente.
Declaración de Ética
Los estudios con participantes humanos fueron revisados y aprobados por la Cuarta Universidad Médica Militar de la Universidad de Beihang. Los pacientes / participantes proporcionaron su consentimiento informado por escrito para participar en este estudio.
Contribuciones de los autores
YS diseñó los experimentos. ZP produjo los resultados y escribió el manuscrito. CD y LZ analizaron e interpretaron los datos. JT e YS realizaron los experimentos, conocieron los datos y los garantes de este estudio. YB, LZ y JT contribuyeron a participar en la recolección de datos y la revisión de la literatura. Todos los autores listados han leído y aprobado el manuscrito final.
Financiación
Esta investigación fue apoyada por la Fundación Nacional de Ciencia del Plan Tecnológico de los Juegos Olímpicos de Invierno de China bajo los números de Subvención 2019YFF0301600 y HJ20191A020135.
Conflicto de Intereses
Los autores declaran que la investigación se realizó en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un conflicto de intereses potencial.
Anderson, C., and Platten, C. R. (2011). La privación del sueño reduce la inhibición y mejora la impulsividad a los estímulos negativos. Behav. Brain Res .. 217:466. doi: 10.1016 / j. bbr.2010.09.020
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Baddeley, A. (2000). El tampón episódico: un nuevo componente de la memoria de trabajo? Trends Cogn. Sci. 4:417. doi: 10.1016 / s1364-6613(00)01538-2
Texto completo cruzado | Google Scholar
Bartel, P., Offermeier, W., Smith, F., and Becker, P. (2004). Atención y memoria de trabajo en anestesistas residentes después del trabajo nocturno: efectos grupales e individuales. Occup. Environ. Mediterráneo. 61, 167–170. doi: 10.1136 / oem.2002.006197
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Casement, M. D., Broussard, J. L., Mullington, J. M., and Press, D. Z. (2006). La contribución del sueño a las mejoras en la velocidad de escaneo de la memoria de trabajo: un estudio de la restricción prolongada del sueño. Biol. Psicol. 72:212. doi: 10.1016 / j. biopsycho.2005.11.002
Resumen de PubMed / Texto completo cruzado / Google Scholar
Chee, M. W. L., and Chuah, L. Y. M. (2008). Comprensión de neuroimagen funcional de cómo el sueño y la privación de sueño afectan la memoria y la cognición. Curr. Opin. Neurol. 21, 417–423. doi: 10.1097/OMA.0b013e3283052cf7
PubMed Abstract / CrossRef Full Text / Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S., and Chee, M. W. L. (2005). Disociación de regiones corticales modulada tanto por la carga de memoria de trabajo como por la privación de sueño y por la privación de sueño sola. Neuroimagen 25, 579-587. doi: 10.1016 / j.neuroimagen.2004.11.029
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Chuah, Y. M. L., Venkatraman, V., Dinges, D. F., and Chee, M. W. L. (2006). La base neuronal de la variabilidad interindividual en la eficiencia inhibitoria después de la privación del sueño. J. Neurocirujano. 26, 7156–7162. doi: 10.1523 / jneurosci.0906-06. 2006
CrossRef Full Text/Google Scholar
Cote, K. A., Milner, C. E., Osip, S.L., Baker, M. L., and Cuthbert, B. P. (2008). Excitación fisiológica y atención durante una semana de restricción continua del sueño. Fisiol. Behav. 95, 353–364. doi: 10.1016 / j. physbeh.2008.06.016
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Cote, K. A., Mondloch, C. J., Sergeeva, V., Taylor, M., and Semplonius, T. (2014). Impacto de la privación total del sueño en el procesamiento neuronal conductual de rostros emocionalmente expresivos. Exp. Brain Res. 232, 1429-1442. doi: 10.1007 / s00221-013-3780-1
Resumen de PubMed / Texto completo cruzado / Google Scholar
Donchin, E., and Fabiani, M. (1991). El uso de potenciales cerebrales relacionados con eventos en el estudio de la memoria: ¿es p300 una medida de la distinción de eventos? Psicol. Facul. Publico. 7, 58–77. doi: 10.1111 / sltb.12055
Resumen de PubMed / Texto completo cruzado / Google Scholar
Drummond, S. P., Brown, G. G., Salamat, J. S., and Gillin, J.C. (2004). El aumento de la dificultad de la tarea facilita la respuesta compensatoria cerebral a la privación total del sueño. Dormir 27, 445-451. doi: 10.1093 / sleep / 27.3.445
Texto completo cruzado/Google Scholar
Drummond, S. P. A., and Brown, G. G. (2001). Los efectos de la privación total del sueño en las respuestas cerebrales al rendimiento cognitivo. Neuropsicofarmacología 25, S68-S73. doi: 10.1016 / S0893-133X (01)00325-6
CrossRef Full Text | Google Scholar
Evans, K. M., and Federmeier, K. D. (2007). La memoria que está a la derecha y la memoria que está a la izquierda: los potenciales relacionados con eventos revelan asimetrías hemisféricas en la codificación y retención de información verbal. Neuropsychologia 45, 1777-1790. doi: 10.1016 / j. neuropsicología.2006.12.014
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M., and Höller, Y. (2007). El componente Visual p2 está relacionado con el bloqueo de fase theta. Neurociencia. Lett. 426, 181–186. doi: 10.1016 / j.neulet.2007.08.062
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Géraldine, R., Desgranges, B., Foret, J., and Eustache, F. (2005). Las relaciones entre los sistemas de memoria y las etapas de sueño. J. Sleep Res. 14, 123-140. doi: 10.1111 / j. 1365-2869. 2005. 00450.x
Resumen de PubMed / Texto completo cruzado / Google Scholar
Gosselin, A., Koninck, J. D., and Campbell, K. B. (2005). Privación total de sueño y procesamiento novedoso: implicaciones para el funcionamiento del lóbulo frontal. Clin. Neurofisiol. 116, 211–222. doi: 10.1016 / j.clinph.2004.07.033
Resumen de PubMed | Texto completo Cruzado/Google Scholar
Harrison, Y., and Horne, J. (1998). La pérdida de sueño perjudica las tareas de lenguaje cortas y novedosas que tienen un enfoque prefrontal. J. Sleep Res. 7, 95-100. doi: 10.1046 / j. 1365-2869. 1998. 00104.x
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J., and Meier, B. (2010). Validez concurrente de la tarea n-back como medida de memoria de trabajo. Memoria 18, 394-412. doi: 10.1080 / 09658211003702171
Resumen de PubMed / Texto completo cruzado/Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., and Chamari, K. (2013). El impacto de la privación parcial del sueño en las variaciones diurnas del rendimiento cognitivo en sujetos entrenados. Proc. Soc. Behav. Sci. 82, 392–396. doi: 10.1016 / j. sbspro.2013.06.281
CrossRef Full Text/Google Scholar
Jennings, J. R., Monk, T. H., and Molen, M. W. V. D. (2003). La privación del sueño influye en algunos, pero no en todos, los procesos de atención supervisora. Psicol. Sci. 14, 473–479. doi: 10.1111/1467-9280. 02456
Resumen de PubMed | Texto completo cruzado/Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L., and Lei, Y. (2015). Recuperación del sueño revierte la inhibición de la respuesta alterada debido a la restricción del sueño: evidencia de un estudio de potenciales relacionados con eventos visuales. PLoS One 10:e0142361. doi: 10.1371 / journal.ponga.0142361
Resumen de PubMed / Texto completo cruzado / Google Scholar
Jones, K., and Harrison, Y. (2001). Función del lóbulo frontal, pérdida de sueño y sueño fragmentado. Medicina para Dormir. Apocalipsis 5: 475. doi: 10.1053 / smrv.2001.0203
Resumen de PubMed / Texto completo cruzado / Google Scholar
Koslowsky, M., and Babkoff, H. (1992). Metaanálisis de la relación entre la privación total de sueño y el rendimiento. Cronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
Resumen de PubMed | Texto completo cruzado/Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A., and Hansenne, M. (2014). El abuso de alcohol y los componentes de erp en tareas activas/no activas utilizando estímulos relacionados con el alcohol: impacto de evitar el alcohol. Int. J. Psychophysiol. 94, 92–99. doi: 10.1016 / j. ijpsycho.2014.08.001
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F., and Huster, R. J. (2019). La privación del sueño afecta de manera diferente a los subcomponentes del control cognitivo. Sueño 42: zsz016. doi: 10.1093 / sleep / zsz016
Resumen PubMed / Texto completo cruzado / Google Scholar
Lavie, P. (2001). Sueño-vigilia como ritmo biológico. Annu. Reverendo Psychol. 52, 277–303. doi: 10.1146 / annurev.psiquiatría.52.1.277
Texto completo cruzado / Google Scholar
Lei, Y., Shao, Y., Wang, L., Zhai, T., and Zou, F. (2015). El acoplamiento de redes cerebrales a gran escala predice los efectos de la privación total del sueño en la capacidad cognitiva. PLoS One 10: e0133959. doi: 10.1371 / journal.ponga.0133959
Resumen de PubMed / Texto completo cruzado / Google Scholar
Li, C., Huang, D., Qi, J., Chang, H., and Meng, Q. (2016). Los déficits de memoria a corto plazo se correlacionan con alteraciones de la conectividad funcional hipocampo-talámico después de la restricción aguda del sueño. Las Imágenes Cerebrales Se Comportan. 11, 954–963. doi: 10.1007 / s11682-016-9570-1
Resumen de PubMed / Texto completo cruzado / Google Scholar
Ma, N., Dinges, D. F., Basner, M., and Rao, H. (2014). Cómo afecta la pérdida total de sueño aguda al cerebro asistente: un meta-análisis de estudios de neuroimagen. Dormir 38, 233. doi: 10.5665/sueño.4404
Resumen de PubMed / Texto completo cruzado / Google Scholar
Miyake, A., and Shah, P. (1999). Modelos de Memoria de Trabajo. Mecanismos de Mantenimiento Activo y Control Ejecutivo. Cambridge: Cambridge University Press.
Google Scholar
Mograss, M. A., Guillem, F., Brazzini-Poisson, V., and Godbout, R. (2009). The effects of total sleep deprivation on recognition memory processes: a study of event-related potential (en inglés). Neurobiol. Aprender. Mem. 91, 343–352. doi: 10.1016 / j. nlm.2009.01.008
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Montplaisir, J. (1981). Depresión y ritmo biológico: efectos terapéuticos de la privación del sueño. L’unión Médicale du Canada 110, 272-276. doi: 10.1620 / tjem.133.481
Resumen de PubMed | Texto completo Cruzado/Google Scholar
Morris, A. M., So, Y., Lee, K. A., Lash, A. A., y Becker, C. E. (1992). El potencial relacionado con eventos del p300. los efectos de la privación del sueño. J. Occupat. Mediterráneo. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F., and Hobson, J. A. (2002). La corteza prefrontal en el sueño. Trends Cogn. Sci. 6, 475–481. doi: 10.1016 / S1364-6613(02)01992-7
Texto completo cruzado | Google Scholar
Ning, M., Dinges, D. F., Mathias, B., and Hengyi, R. (2014). Cómo afecta la pérdida total de sueño aguda al cerebro asistente: un meta-análisis de estudios de neuroimagen. Dormir 38, 233-240. doi: 10.5665/sueño.4404
Resumen de PubMed | Texto completo cruzado/Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, A. R., and Bullmore, E. (2005). Paradigma de memoria de trabajo N-back: un meta-análisis de estudios de neuroimagen funcional normativa. Zumbido. Mapa Cerebral. 25, 46–59. doi: 10.1002 / hbm.20131
Resumen de PubMed / Texto completo cruzado / Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S., and Kumar, S. (2010). Los correlatos electrofisiológicos de la cognición mejoran con la siesta durante la privación del sueño. EUR. J. Appl. Fisiol. 108, 549–556. doi: 10.1007 / s00421-009-1222-3
Resumen de PubMed / Texto completo cruzado / Google Scholar
Ratcliff, R., and Van Dongen, H. (2018). Los efectos de la privación de sueño sobre el ítem y la memoria de reconocimiento asociativo. J. Exp. Psicol. 44, 193–208. doi: 10.1037 / xlm0000452
Resumen PubMed / Texto completo cruzado / Google Académico
Raymond, C. A. (1988). El cambio de trabajo y los ciclos de sueño se están convirtiendo en otro problema de salud pública. JAMA J. Am. Mediterráneo. Assoc. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef Full Text / Google Scholar
Saito, H., Yamazaki, H., Matsuoka, H., Matsumoto, K., and Sato, M. (2001). Potencial relacionado con eventos visuales en demencia leve del tipo de alzheimer. Psiquiatría Clin. Neurociencia. 55, 365–371. doi: 10.1046 / j. 1440-1819. 2001. 00876.x
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Schacht, A., Werheid, K., and Sommer, W. (2008). La valoración de la belleza facial es rápida pero no obligatoria. Cogn. Afectar. Behav. Neurociencia. 8, 132–142. doi: 10.3758 / cabn.8.2.132
Resumen de PubMed / Texto completo cruzado / Google Scholar
Scott, J. P. R., Mcnaughton, L. R., and Polman, R. C. J. (2006). Efectos de la privación de sueño y el ejercicio sobre el rendimiento cognitivo, motor y el estado de ánimo. Fisiol. Behav. 87:408. doi: 10.1016 / j. physbeh.2005.11.009
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Smallwood, J., Brown, K., Baird, B., and Schooler, J. W. (2011). Cooperación entre la red de modo predeterminado y la red frontal-parietal en la producción de un tren de pensamiento interno. Brain Res. 1428, 60-70. doi: 10.1016 / j. brainres.2011.03.072
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Smith, M. E., Mcevoy, L. K., and Gevins, A. (2002). El impacto de la pérdida moderada de sueño en las señales neurofisiológicas durante la realización de tareas de memoria de trabajo. Dormir 25, 784-794. doi: 10.1093 / sueño / 25.7.56
CrossRef Full Text / Google Scholar
Thomas, M. (2003). Base neuronal del estado de alerta y deterioro del rendimiento cognitivo durante la somnolencia ii. efectos de 48 y 72 h de privación de sueño en la actividad cerebral regional humana despierta. Thalam. Relat. Syst. 2, 199–229. doi: 10.1016 / S1472-9288(03)00020-7
Texto completo cruzado | Google Scholar
Van Dongen, H. (2004). El costo acumulado de la vigilia adicional: efectos de respuesta de dosis sobre las funciones neuroconductuales y la fisiología del sueño de la privación crónica del sueño y la privación total del sueño. Dormir 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef Full Text/Google Scholar
Verweij, I. M., Romeijn, N., Smit, D. J., and Giovanni, P. (2014). La falta de sueño conduce a una pérdida de conectividad funcional en las regiones frontales del cerebro. Neurociencia de BMC. 15:88. doi: 10.1186/1471-2202-15-88
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Wen, X., Liu, Y., Yao, L., and Ding, M. (2013). Regulación de arriba hacia abajo de la actividad del modo predeterminado en la atención visual espacial. J. Neurocirujano. 33, 6444–6453. doi: 10.1523 / JNEUROSCI.4939-12.2013
Texto completo cruzado | Google Scholar
Wiggins, E., Malayka, M., and Kendra, G. (2018). la privación de sueño de 24 horas afecta la modulación atencional temprana del procesamiento neural: un estudio de potencial cerebral relacionado con eventos. Neurociencia. Lett. 677, 32–36. doi: 10.1016 / j.neulet.2018.04.022
Resumen de PubMed / Texto completo cruzado / Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y., and Zhou, Q. (2019). Disminución de la información reemplazo de la memoria de trabajo después de la privación del sueño: evidencia de un estudio potencial relacionado con eventos. Delantero. Neurociencia. 13:408. doi: 10.3389 / fnins.2019.00408
Texto completo cruzado / Google Scholar
Zhang, Q., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z., et al. (2014). Un estudio visual erp de la inhibición de impulsos después de una siesta inducida por zaleplon después de la privación del sueño. PLoS One 9: e95653. doi: 10.1371 / journal.ponga.0095653
Resumen de PubMed / Texto completo de CrossRef / Google Scholar
Leave a Reply