Estantería
A pesar de su ubicación periférica, la retina o porción neural del ojo, es en realidad parte del sistema nervioso central. Durante el desarrollo, la retina se forma como una salida del diencéfalo, llamada vesícula óptica, que sufre invaginación para formar la copa óptica (Figura 11.3; ver también Capítulo 22). La pared interna de la copa óptica da lugar a la retina, mientras que la pared externa da lugar al epitelio pigmentario. Este epitelio es una estructura que contiene melanina y reduce la retrodispersión de la luz que entra en el ojo; también desempeña un papel crítico en el mantenimiento de los fotorreceptores, renovando los fotopigmentos y fagocitando los discos fotorreceptores, cuyo recambio a una alta velocidad es esencial para la visión.

Figura 11.3
el Desarrollo del ojo humano. (A) La retina se desarrolla como una salida del tubo neural, llamada vesícula óptica. B) La vesícula óptica se invagina para formar la copa óptica. (C, D)La pared interna de la copa óptica se convierte en la retina neural, mientras que la (más…)
Consistente con su estado como una parte completa del sistema nervioso central, la retina comprende circuitos neuronales complejos que convierten la actividad eléctrica graduada de los fotorreceptores en potenciales de acción que viajan al cerebro a través de axones en el nervio óptico. Aunque tiene los mismos tipos de elementos funcionales y neurotransmisores que se encuentran en otras partes del sistema nervioso central, la retina comprende solo unas pocas clases de neuronas, y estas están dispuestas de una manera que ha sido menos difícil de desentrañar que los circuitos en otras áreas del cerebro. Hay cinco tipos de neuronas en la retina: fotorreceptores, células bipolares, células ganglionares, células horizontales y células amacrinas. Los cuerpos celulares y los procesos de estas neuronas se apilan en cinco capas alternas, con los cuerpos celulares ubicados en las capas de células ganglionares, nucleares internas y nucleares externas, y los procesos y contactos sinápticos ubicados en las capas plexiformes internas y externas (Figura 11.4). Una cadena directa de tres neuronas-célula fotorreceptora a célula bipolar a célula ganglionar—es la principal ruta de flujo de información desde los fotorreceptores hasta el nervio óptico.
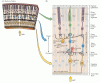
Figura 11.4
Estructura de la retina. (A) Sección de la retina que muestra la disposición general de las capas retinianas. (B) Diagrama de los circuitos básicos de la retina. Una cadena de tres neuronas-fotorreceptor, célula bipolar y célula ganglionar—proporciona la más directa (más…)
Hay dos tipos de elementos sensibles a la luz en la retina: bastones y conos. Ambos tipos de fotorreceptores tienen un segmento externo que está compuesto de discos membranosos que contienen fotopigmento y se encuentra adyacente a la capa epitelial pigmentaria, y un segmento interno que contiene el núcleo celular y da lugar a terminales sinápticos que entran en contacto con células bipolares u horizontales. La absorción de luz por el fotopigmento en el segmento externo de los fotorreceptores inicia una cascada de eventos que cambia el potencial de membrana del receptor y, por lo tanto, la cantidad de neurotransmisor liberada por el fotorreceptor sinapta en las células que entran en contacto. Las sinapsis entre los terminales fotorreceptores y las células bipolares (y las células horizontales) ocurren en la capa plexiforme externa; más específicamente, los cuerpos celulares de los fotorreceptores constituyen la capa nuclear externa, mientras que los cuerpos celulares de las células bipolares se encuentran en la capa nuclear interna. Los procesos axonales cortos de las células bipolares hacen contactos sinápticos a su vez en los procesos dendríticos de las células ganglionares en la capa plexiforme interna. Los axones mucho más grandes de las células ganglionares forman el nervio óptico y transportan información sobre la estimulación retiniana al resto del sistema nervioso central.
Los otros dos tipos de neuronas en la retina, las células horizontales y las células amacrinas, tienen sus cuerpos celulares en la capa nuclear interna y son los principales responsables de las interacciones laterales dentro de la retina. Estas interacciones laterales entre receptores, células horizontales y células bipolares en la capa plexiforme externa son en gran parte responsables de la sensibilidad del sistema visual al contraste de luminancia en un amplio rango de intensidades de luz. Los procesos de las células amacrinas, que se extienden lateralmente en la capa plexiforme interna, son postsinápticos a las terminales celulares bipolares y presinápticos a las dendritas de las células ganglionares (véase la Figura 11.4). Los procesos de las células horizontales se ramifican en la capa plexiforme externa. Varias subclases de células amacrinas que hacen distintas contribuciones a la función visual. Una clase de células amacrinas, por ejemplo, juega un papel importante en la transformación de las respuestas persistentes de las células bipolares a la luz en las breves respuestas transitorias exhibidas por algunos tipos de células ganglionares. Otro tipo sirve como paso obligatorio en la vía que transmite información de los fotorreceptores de bastones a las células ganglionares de la retina. La variedad de subtipos de células amacrinas ilustra la regla más general de que, aunque solo hay cinco tipos básicos de células retinianas, puede haber una diversidad considerable dentro de un tipo de célula dado. Esta diversidad es la base de vías que transmiten diferentes tipos de información a los objetivos centrales de manera paralela.
A primera vista, la disposición espacial de las capas retinianas parece contradictoria, ya que los rayos de luz deben pasar a través de los elementos no sensibles a la luz de la retina (¡y la vasculatura retiniana!) antes de alcanzar los segmentos exteriores de los fotorreceptores, donde se absorben los fotones (véase la Figura 11.4). La razón de esta característica curiosa de la organización retiniana radica en la relación especial que existe entre los segmentos externos de los fotorreceptores y el epitelio pigmentario. Los segmentos exteriores contienen discos membranosos que albergan el fotopigmento sensible a la luz y otras proteínas involucradas en el proceso de transducción. Estos discos se forman cerca del segmento interno del fotorreceptor y se mueven hacia la punta del segmento externo, donde se desprenden. El epitelio pigmentario desempeña un papel esencial en la eliminación de los discos receptores gastados; esta tarea no es pequeña, ya que todos los discos en los segmentos externos se reemplazan cada 12 días. Además, el epitelio pigmentario contiene la maquinaria bioquímica necesaria para regenerar el fotopigmento moléculas después de haber sido expuestas a la luz. Es presumiblemente las demandas del ciclo de vida del disco fotorreceptor y el reciclaje de fotopigmentos lo que explica por qué las varillas y los conos se encuentran en la capa más externa en lugar de en la capa más interna de la retina. Las alteraciones en las relaciones normales entre el epitelio pigmentario y los fotorreceptores retinianos, como los que se producen en la retinitis pigmentosa, tienen consecuencias graves para la visión (Recuadro B).
Box B
Retinitis Pigmentosa.
Leave a Reply