Enlace peptídico
La deslocalización significativa del par solitario de electrones en el átomo de nitrógeno le da al grupo un carácter de doble enlace parcial. El doble enlace parcial hace que el grupo amida sea plano, que ocurre en los isómeros cis o trans. En el estado desplegado de las proteínas, los grupos de péptidos son libres de isomerizarse y adoptar ambos isómeros; sin embargo, en el estado plegado, solo se adopta un solo isómero en cada posición (con raras excepciones). La forma trans se prefiere abrumadoramente en la mayoría de los enlaces peptídicos (aproximadamente una proporción de 1000:1 en las poblaciones trans:cis). Sin embargo, los grupos de péptidos X-Pro tienden a tener una relación de aproximadamente 30:1, presumiblemente debido a la simetría entre C α {\displaystyle \mathrm {C^{\alpha }} }

y C δ {\displaystyle \mathrm {C^{\delta }} }

los átomos de prolina hacen que los isómeros cis y trans sean casi iguales en energía (ver figura, a continuación).
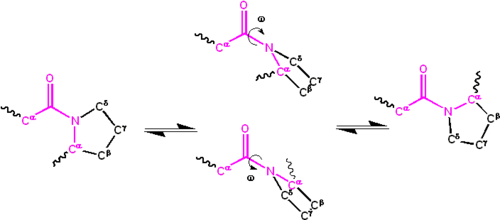
El ángulo diedro asociados con el péptido grupo (definido por los cuatro átomos de C α − C ‘ − N − C α {\displaystyle C^{\alpha }-C^{\prime }-N-C^{\alpha }}

) se denota ω {\displaystyle \omega }

; ω = 0 ∘ {\displaystyle \omega =0^{\circ }}

para el isómero cis (synperiplanar conformación) y ω = 180 ∘ {\displaystyle \omega =180^{\circ }}

para el isómero trans (antiperiplanar conformación). Los grupos de amidas pueden isomerizarse sobre el enlace C ‘- N entre las formas cis y trans, aunque lentamente (τ {{\displaystyle \ tau \ sim}

20 segundos a temperatura ambiente). Los estados de transición ω = ± 90 ∘ {\displaystyle \omega =\pm 90^{\circ }}

requiere que el parcial de doble enlace roto, por lo que la energía de activación es de aproximadamente 80 kilojoule/mol (20 kcal/mol). Sin embargo, la energía de activación puede reducirse (y la isomerización catalizada) mediante cambios que favorecen la forma de enlace único, como colocar el grupo de péptidos en un entorno hidrofóbico o donar un enlace de hidrógeno al átomo de nitrógeno de un grupo de péptidos X-Pro. Ambos mecanismos para reducir la energía de activación se han observado en las isomerasas peptidil prolil (PPIasas), que son enzimas naturales que catalizan la isomerización cis-trans de los enlaces peptídicos X-Pro.
El plegamiento de proteínas conformacionales suele ser mucho más rápido (típicamente 10-100 ms) que la isomerización cis-trans (10-100 s). Un isómero no nativo de algunos grupos de péptidos puede interrumpir significativamente el plegado conformacional, ya sea ralentizándolo o evitando que incluso ocurra hasta que se alcance el isómero nativo. Sin embargo, no todos los grupos de péptidos tienen el mismo efecto en el plegado; es posible que los isómeros no nativos de otros grupos de péptidos no afecten en absoluto al plegado.
Leave a Reply