gränser i neurovetenskap
introduktion
med samhällets framsteg och förändringar i arbetsrytmen lider ett ökande antal människor av sömnbrist. Sömnbrist skadar inte bara individens fysiska och psykiska hälsa utan påverkar också allvarligt arbetsprestanda, vilket orsakar arbetsfel och till och med olyckor. Att förstå mekanismen för sömnbrist som påverkar kognitiv funktion är därför av stor betydelse för att effektivt förhindra effekterna av sömnbrist.
tidigare studier har visat att sömnbrist kan orsaka en serie förändringar i individens humör, kognitiva förmåga, arbetsprestanda och immunfunktion (Choo et al., 2005). Bristen på sömn stör kroppens cirkulation och påverkar individens kognitiva och emotionella förmågor (Raymond, 1988). Flera studier har visat att sömnbrist försämrar responshämning (Harrison och Horne, 1998; Muzur et al., 2002; Jennings et al., 2003). Till exempel, efter 36 timmar av sömnbrist minskade individens förmåga att undertrycka negativa stimuli (Chuah et al., 2006). Neuroimaging studier har föreslagit att sömnbrist minskar individens låga nivå av visuell bearbetningsförmåga (Anderson och Platten, 2011; Ning et al., 2014). Dessutom försämrar sömnbrist hippocampus och kan påverka minnet genom att förstöra synaptisk plasticitet (Cote et al., 2014). Thomas (2003) har indikerat att brist på sömn minskade cerebralt blodflöde och metabolisk hastighet i thalamus, prefrontal cortex och parietal cortex (g Bikraldine et al., 2005). Jarraya och kollegor fann att partiell sömnbrist påverkade neuropsykologiska funktioner som verbalt omedelbart minne, uppmärksamhet och vakenhet (Thomas, 2003). Dessutom har vissa studier visat att de kumulativa effekterna av partiell sömnbrist kan allvarligt försämra kognitiv funktion och beteende (Van Dongen, 2004; Scott et al., 2006; Jarraya et al., 2013).
arbetsminne är ett system som används för att lagra och bearbeta information och som är en kognitiv funktion med begränsad kapacitet (Bartel et al., 2004). Dessutom kan informationen som lagras i arbetsminnessystemet ändras från korttidsminne till långtidsminne genom återställning och andra minnesmetoder. Arbetsminne är övergången mellan kortvariga och långsiktiga minnessystem, vilket är mycket avgörande för mänsklig meddelandebehandling (Miyake och Shah, 1999). Det ger ett tillfälligt lagringsutrymme och de resurser som behövs för att bearbeta information, såsom röstförståelse, resonemang och lärande. Sömnbrist har visat sig påverka arbetsminnet först.
tidigare studier har använt N-back arbetsminnesparadigmet hos deltagare som genomgick sömnbrist och fann att brist på sömn inducerar en minskning av metabolisk aktivitet i hjärnans regionala nätverk, vilket huvudsakligen sker informationsbehandling och reaktionshämning (Baddeley, 2000; Zhang et al., 2019). Nedsatt arbetsminne efter sömnbrist är relaterat till aktiveringen av standardnätverket i uppgifter (Chee och Chuah, 2008), vilket kan vara relaterat till thalamusens viktiga roll i kortikal vakenhet. Till exempel ökade sömnbrist sambandet mellan hippocampus, thalamus och standardnätverk, vilket ofta åtföljdes av högre subjektiv dåsighet och sämre prestanda i arbetsminnet (Lei et al., 2015; Li et al., 2016). Studier om sömnbrist identi?ed att ökad latens och minskad amplitud av P3-komponenten var associerad med långvarig nykterhet (Morris et al., 1992; Jones och Harrison, 2001; Panjwani et al., 2010). Minskningen i P3-vågen kan återspegla en minskning av deltagarnas uppmärksamhet och en minskning av urskiljningen av målstimuli (Koslowsky och Babkoff, 1992).
men få studier har gett elektrofysiologiska bevis för nedsatt arbetsminne efter sömnbrist. N-back-uppgiften anses vara en vanlig metod för att bedöma arbetsminnet (Owen et al., 2005; Jaeggi et al., 2010). Zhang et al. utformad en två-back uttal arbetsminne uppgift att utforska den minskade meddelande alternativ arbetsminne under sömnbrist, men få studier har använt olika typer av arbetsminne uppgifter i ett enda experiment. I den aktuella studien utformade vi olika typer av arbetsminneuppgifter (uttal arbetsminne, rumsligt arbetsminne och objektarbetsminne) för att utforska försämringen av kognitiv funktion av TSD och inspelad deltagares EEG-data vid 2 tidpunkter (baslinje och 36 h-TSD). Alla uppgifter antog ett 2-back paradigm. Denna studie utvärderade förändringarna i N2-P3-vågen relaterad till arbetsminne under TSD och analyserade de temporära egenskaperna hos effekterna av sömnbrist på arbetsminnet. Våra resultat ger experimentella bevis för effekterna av sömnbrist på kognitiv funktion.
material och metoder
deltagare
sexton unga, friska, högerhänta manliga studenter deltog i denna studie. Vi rekryterade deltagare genom reklam på campus. Deltagarna hade alla goda sömnvanor (PSQI<5). Alla deltagare var mellan 21 och 28 år med en medelålder på 23 år, och ingen av deltagarna hade någon psykisk eller fysisk sjukdom. Alla deltagare hade normal syn eller korrigerad syn över 1.0 och intelligenspoäng >110 på Raven-testet. Före experimentet förklarade experimenten proceduren och pekar på uppmärksamhet åt deltagarna för att se till att de var bekanta med metoden och proceduren. Under de 2 veckorna före experimentet sov deltagarna regelbundet i 7-9 timmar per dag, utan att röka, dricka kaffe, dricka alkohol eller konsumera något läkemedel i 2 dagar före experimentet. Före experimentet gav alla deltagare skriftligt informerat samtycke. Det experimentella systemet godkändes av Etikutskottet vid det fjärde militära medicinska universitetet och Beihang University.
experimentell Design
tre typer av arbetsminneuppgifter presenterades för alla deltagare. De var två-back uttal arbetsminne uppgift (se Figur 1), två-back rumsliga arbetsminne uppgift (se Figur 2), och två-back objekt arbetsminne uppgift (se Figur 3). De stimulerande materialen i uppgifterna var 15 skiftlägeskänsliga engelska bokstäver som utesluter de med liknande bokstäver, såsom L/l, M/m; små svarta rutor; respektive 12 geometriska figurer. Alla material visades i svart färg på en vit bakgrund, med en ungefärlig visuell vinkel på 1,5 1,5 1,5 (bredd: 2,0 cm, Höjd: 2,0 cm) subtending. 122 spår bestod i varje uppgift och i varje spår presenterades målstimulansen för 400 ms två spår efter presentationen av objektiv stimulans, med 1,600 ms stimulus start asynchrony time (SOA) som präglades av en vit ”+.”Deltagarna ombads att klicka på vänster musknapp när målet och den objektiva stimulansen var desamma (”matchning”), medan du klickar på höger musknapp när de inte var (”felaktig matchning”). Matchnings-eller inte-villkoret presenterades i en pseudorandomordning med ett förhållande 1:1.
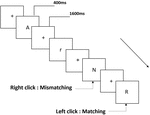
Figur 1. Schematiskt diagram över uttalet arbetsminne uppgift.
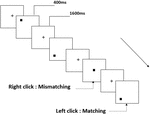
Figur 2. Schematiskt diagram över det rumsliga arbetsminnesuppgiften.
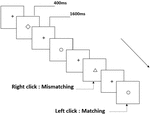
Figur 3. Schematiskt diagram över objektet arbetsminne uppgift.
experimentella procedurer
före experimentet instruerades deltagarna om den experimentella uppgiften. De informerades om att öva de tre typerna av arbetsminneuppgifter tills en noggrannhet på 90% uppnåddes. Deltagarna besökte laboratoriet dagen före experimentet och sov i laboratoriet den natten. De två partnerdeltagarna utförde experimenten samtidigt. Tre typer av arbetsminneuppgifter utfördes klockan 7: 30 till 8:30 nästa morgon med samtidig elektroencefalogram (EEG) inspelning (baslinje). Den andra EEG-inspelningen (36 h-TSD) genomfördes efter en 36-h-period under vilken deltagarna inte fick sova. Under hela experimenttiden förbjöds central hämning och stimulerande läkemedel. Deltagarna åtföljdes, observerades och påminde av vårdpersonal för att hålla dem vakna under hela TSD-sessionen.
EEG Recordings
en kontinuerlig scalp EEG spelades in med elektrodkapslar placerade på 64 platser med 10-20-systemet med en SynAmps2-förstärkare. De bilaterala mastoiderna (A1 och A2) användes som referens och pannan användes som mark. EEG registrerades vid 1000 Hz, och impedansen för alla kanaler bibehölls under 5 k KB. Fyra ytterligare elektroder placerades ovanför och under höger och vänster ögon för att registrera ett bipolärt vertikalt och horisontellt elektrookulogram.
dataanalys av beteendeexperiment
på grund av tekniska fel raderades två fall medan andra 14 fall inkluderades i följande statistiska analys. Beteendedata inkluderade den genomsnittliga reaktionstiden, rätt hastighet och rätt antal per tidsenhet. Beteendedata i baslinjen och 36 h-TSD-tillstånd registrerades för analys. Analyserna drivs av IBM SPSS (V22.2), där metoden för upprepad måttanalys av varians (ANOVA) med växthus-Geisser var Bonferroni post-hoc-analys lanserades. De statistiska resultaten presenterades som medel-och standardavvikelse (SD).
EEG-dataanalys
Scan 4.3-programmet användes för att analysera EEG-data, där EEG-artefakterna i ögonrörelsen korrigerades med okulär artefaktreduceringsmetod. Epoker som sträcker sig från -100 till 800 ms av de kontinuerliga EEG-data extraherades och filtrerades av ett bandpassfilter från 0,5 till 30 Hz med frekvenslutningen på 24 dB/oct. Försök i vilken spänning har överskridit ± 100 µV förkastades och den baslinje som har korrigerats till en genomsnittlig amplitud av 100 ms. EEG komponenter var i genomsnitt och beräknas med endast det rättade svar. ERP-komponenterna P2 (100-250 ms), N2 (150-350 ms) och P3 (250-450 ms) i stimulansstudierna identifierades och kvantifierades. De stora genomsnittliga toppamplituderna och latenserna för N2-och P3-komponenterna beräknades separat vid F3, Fz, F4, C3, Cz, C4, P3, Pz och P4, och P2-komponenten beräknades vid F3, Fz, F4, C3, Cz och C4 (Casement et al., 2006; Verweij et al., 2014).
upprepade åtgärder ANOVA användes för alla ERP-resultat. Huvudeffekterna och interaktionerna mellan sömntillstånd (baslinje och 36 h-TSD), uppgifter (uttal arbetsminne, rumsligt arbetsminne och objektarbetsminne), regioner (frontal, central och parietal; P2-komponenten analyserades endast på de främre och centrala regionerna) och platser (vänster, mitten och höger) analyserades statistiskt med hjälp av upprepade åtgärder ANOVA, som inkluderade växthus-Geisser-korrigeringar för icke-sfärisk och Bonferroni post-hoc-test.
resultat
Beteendeprestanda
resultaten av beteendeexperimenten visas i Tabell 1. Den genomsnittliga reaktionstiden var längre i 36 h-TSD-tillståndet än vid baslinjen med en trend att öka men utan signifikanta skillnader. ANOVA avslöjade att den korrekta hastigheten för uppgiften var signifikant olika mellan baslinjen och 36 h-TSD . Det korrekta antalet per tidsenhet visade en signifikant huvudeffekt av tiden under 36 h-TSD .

tabell 1. Prestandadata (medelvärde för 2-back-uppgiften vid baslinjen och efter 36 h-TSD.
Amplitud
jämfört med baslinjen observerades en signifikant minskning i amplituden för P3 och en signifikant ökning observerades i amplituden för P2 efter TSD. Även om N2-amplituden minskade efter 36 h TSD, nådde skillnaden inte statistisk signifikans (Tabell 2).

tabell 2. Grand-Genomsnittlig toppamplitud för P2 -, N2-och P3-komponenterna i rätt svarstillstånd över flera elektrodställen vid baslinjen och efter 36 h-TSD.
signifikanta huvudeffekter av regioner och platser på P2-amplituden hittades under TSD-tillståndet. Under TSD uppträdde den maximala amplituden för P2 i frontalområdet (Figur 4). Dessutom var skillnaderna i P2-amplituder i olika regioner (frontal vs central) mer signifikanta i de bilaterala elektroderna (vänster: P = 0,001; höger: P = 0,000) än i mellanelektroderna (Figur 4). En signifikant huvudeffekt av regionen och platsen på N2 avslöjade att N2-amplituden var mer negativ i fronten än i den centrala regionen (P = 0,008, Figur 4) och var mindre till höger än på vänster sida (P = 0,011, figurer 5A2,B2). En huvudeffekt av site på P3-amplituden observerades . Amplituden för P3 var mer positiv i mitten än på vänster sida (P = 0,009, Figur 4). En signifikant interaktionseffekt mellan tid och region observerades för P3-amplituden . Under TSD var minskningen av P3-amplituden mer signifikant i de främre och centrala regionerna än i parietalområdet (P = 0,005; P = 0,003) (figur 5C3). Inga andra huvudeffekter eller interaktionseffekter nådde statistisk signifikans.
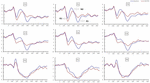
Figur 4. ERP-amplitud vid baslinjen och 36 h-TSD för rätt svarstillstånd för arbetsminnesuppgiften. Kanalerna beställs från vänster till höger och uppifrån och ner enligt följande: F3, Fz, F4, C3, Cz, C4, P3, Pz och P4. Jämfört med baslinjen förlängdes latenserna för N2-P3-komponenterna och amplituderna för N2-P3 minskade efter 36 h-TSD.
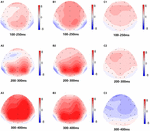
Figur 5. Topografisk karta över Rätt svar i arbetsminnesuppgiften under olika sömnförhållanden (A1-C3). (A1) P2, 100-250 ms, vid baslinjen. (A2) N2, 200-300 ms, vid baslinjen. (A3) P3, 300-400 ms, vid baslinjen. (B1) P2, 100-250 ms, vid 36 h-TSD. (B2) N2, 200-300 ms, för 36 h-TSD. (B3) P3, 300-400 ms, för 36 h-TSD. (C1) P2, 100-250 ms, 36 h-TSD med subtraherad baslinje. (C2) N2, 200-300 ms, 36 h-TSD med subtraherad baslinje. (C3) P3, 300-400 ms, 36 h-TSD med subtraherad baslinje.
latens
latenserna för N2 och P2 förlängdes signifikant efter TSD. Även om P3-latensen förlängdes efter 36 timmar av TSD, nådde skillnaden inte statistisk signifikans (tabell 3).

tabell 3. Grand-Genomsnittlig topplatens för P2 -, N2-och P3-komponenterna i rätt svarstillstånd över flera elektrodplatser vid baslinjen och efter 36 h-TSD.
den signifikanta huvudeffekten av regionen på N2 och P3 avslöjade att latensen för N2-P3-komponenterna var kortare i parietalområdet än i frontalområdet (P = 0,002; P = 0,000) och centralregionen (P = 0,000; P = 0,000) (Figur 4). Latensen för P3-vågen var betydligt längre på vänster sida än på höger sida (Figur 4).
inga andra huvudeffekter eller interaktionseffekter nådde statistisk signifikans.
amplituderna och latenserna N2, P2 och P3 som framkallades vid de nio elektrodställena presenteras i Figur 4. Den topografiska kartan över det korrekta svaret i arbetsminnesuppgiften under olika sömnförhållanden (baslinje, 36 h-TSD och skillnaden mellan de två förhållandena) presenteras i Figur 5.
diskussion
i denna studie rapporterade vi påverkan av 36 timmars sömnbrist på arbetsminnet, som kombinerar beteendedata i två sömntillstånd (baslinje och 36 h-TSD) med samtidiga EEG-inspelningar. Analysen av resultaten indikerade att förändringarna i beteendedata i enlighet med nedsatt arbetsminne efter 36 h TSD: en ökning av den genomsnittliga reaktionstiden för de kognitiva uppgifterna och en minskning av noggrannheten.
sömnbrist försämrade individens kontroll av uppmärksamhetsresurser. Även om individer försökte upprätthålla vakenhet och arbetsprestanda, inklusive reaktionstid och korrekt hastighet, under sömnbrist, påverkades informationsbehandlingskapaciteten för deras arbetsminne fortfarande på grund av minskningen av hastigheten för bearbetning av information (Casement et al., 2006; Wiggins et al., 2018). I denna studie mättes N2-och P3-vågorna relaterade till arbetsminnet för att visa en ökning av latens och en minskning av amplituden efter sömnbrist jämfört med baslinjeläsningarna. Studier har visat att sömnbrist leder till en kontinuerlig nedgång i uppmärksamhet, och fenomenet minskad P3-Amplitud indikerar att individers top-down kontroll av kognition gradvis kollapsar. Sömnbrist har en mer negativ effekt på kognitiva funktioner, särskilt de som är beroende av mentala eller kognitioner (Kusztor et al., 2019).
P3-komponenten återspeglar utplaceringen av uppmärksamhetsresurser, och latensen för P3 ses allmänt som tidsfönstret för stimulanskategorisering och utvärdering. Minskningen i P3-vågamplituden bekräftade också att beslutsfattandet i matchningssvaret efter TSD hade skadats i viss utsträckning (Gosselin et al., 2005). Studier har föreslagit att sömnbrist kan påverka informationsbehandlingsstadiet i arbetsminnet. I denna studie stödde resultatindikatorerna också slutsatsen att svarstiden för målstimulansen ökades och att latensen för P3-vågen förlängdes (Cote et al., 2008). Det spekulerades i att effekten av sömnbrist på P3-komponenter också kan äga rum på grund av att de inte svarade på informationsändring, vilket överensstämmer med tidigare slutsatser om att P3-komponenterna är relaterade till uppdatering av arbetsminneinnehåll (Donchin och Fabiani, 1991).
tidigare studier har betraktat N2-komponenten som ett elektrofysiologiskt index som återspeglar individens förmåga att undertrycka svaret (Kreusch et al., 2014). Efter sömnbrist indikerar den långvariga latensen hos Nogo-N2-komponenten att individens förmåga att undertrycka svaret försämras (Jin et al., 2015). Den minskade amplituden och långvariga latensen hos N2-komponenten relaterad till uttal arbetsminne efter sömnbrist avslöjar att sömnbrist försämrar informationsbehandlingen av uttal arbetsminne (Zhang et al., 2019). N2-komponenten anses generellt återspegla hjärnans selektiva uppmärksamhet och bearbetning av känslomässiga stimuli eller signaler (Schacht et al., 2008) och är en endogen komponent relaterad till individens mentala tillstånd, uppmärksamhet och grad av uppmärksamhet. I denna studie fann vi att latensen för N2-komponenten var signifikant förlängd, men amplituden visade endast en nedåtgående trend. Enligt tidigare studier återspeglade långvarig N2-latens en ökning av svarstiden efter sömnbegränsning (Zhang et al., 2014). Upptäckten att N2-amplituden inte signifikant förändrades kan dock ha bero på cerebrala kompensationssvar (Drummond och Brown, 2001). När det gäller begränsade kognitiva resurser fanns det en kompensationsmekanism för att återställa nedsatt kognitiv funktion (Jin et al., 2015).
enligt hårbottentopografin är förändringarna i N2-P3-komponenterna relaterade till sömnbrist mer uppenbara i frontalområdet. Frontoparietal control (FPC) spelar en viktig roll i kognitiv kontroll. Studier har visat att FPC kan kringgå top-down kognitiv kontroll, vilket gör det möjligt för individer att fokusera på information relaterad till målet samtidigt som man undertrycker information som inte är relaterad till målet (Smallwood et al., 2011; Wen et al., 2013). FPC är viktigt för informationslagring och informationsbehandling i arbetsminnet, och graden av aktivering av FPC efter sömnbrist reducerades jämfört med en normal sömngrupp (Ma et al., 2014). Även om EEG-resultaten inte återspeglade förändringarna i specifika hjärnregioner i detalj, återspeglade det intuitivt effekten av TSD på retention och bearbetning av arbetsminnesinformation.
Även om den exakta kognitiva processen som P2-komponenten ligger till grund för fortfarande diskuteras allmänt, som en bred definition, återspeglar P2-komponenten processen med uppmärksamhet och visuell bearbetning och anses allmänt vara relaterad till selektiv uppmärksamhet och arbetsminne, vilket återspeglar den tidiga bedömningen av den perceptuella processen (Saito et al., 2001). I denna studie fann vi en signifikant ökning av P2-vågamplituden efter sömnbrist. Studier har rapporterat att P2-vågor som kan vara en del av det tidiga kognitiva matchningssystemet för meddelandebehandling och kan jämföra sensoriska ingångar till lagrat minne (Freunberger et al., 2007) är känsliga för förändringar i uppdragsuppmärksamhet och arbetsminneskrav (Smith et al., 2002). Funktionell kompensation är en av de mänskliga hjärnans unika funktioner och en viktig faktor för att upprätthålla kognitiv funktion. Överdriven aktivering av den dorsolaterala prefrontala cortexen (DLPFC) efter sömnbrist indikerar att när hjärnresurserna minskar verkar DLPFC ha en kompensationsfunktion (Drummond et al., 2004; Choo et al., 2005). Därför spekulerar vi i att den signifikanta ökningen av P2-amplitud som observerats i denna studie kan bero på funktionell kompensation där individer verkar upprätthålla normal kognitiv funktion efter sömnbrist. Även om ett stort antal studier har använt ERP-teknik för att utforska effekten av sömnbrist på kognitiva funktioner, har tidiga komponenter som N1 och P2 inte studerats systematiskt, och resultaten är inkonsekventa (Evans och Federmeier, 2007; Wiggins et al., 2018; Zhang et al., 2019). Det finns få undersökningar som undersöker förändringen av P2-komponenten under sömnbrist (Mograss et al., 2009). Därför måste effekterna av sömnbrist på tidiga komponenter i ERP, såsom P2, fortfarande studeras och undersökas ytterligare.
i detta experiment använde vi 2-back-modellen för att designa uttal, rumsliga och objektarbetsminneuppgifter och undersökte försämringen av arbetsminnet efter 36 h TSD. Jämfört med tidigare studier som endast fokuserade på effekterna av sömnbrist på en specifik typ av information, såsom uttal arbetsminne eller specifik kognitiv funktion, såsom responshämning, har vi beaktat innehållet i arbetsminnesmodellen och analyserat effekterna av sömnbrist på arbetsminnet.
studien har dock vissa begränsningar. Först använde vi bara 2-back-uppgiften och misslyckades med att jämföra deltagarnas prestanda i arbetsminnesuppgifter med olika svårigheter. Därför finns det begränsningar för att förklara och dra slutsatser om förändringar i arbetsbelastningen. För det andra användes endast manliga volontärer i studien, och slutsatserna måste bedömas när de utvidgas till kvinnliga volontärer. På grund av det begränsade antalet deltagare fann vi bara att amplituden för N2-vågen hade en nedåtgående trend och att P3-vågens latens hade en långvarig trend. Stabila resultat kan erhållas efter att antalet deltagare har ökat. För det tredje kan kombination av vår procedur med fMRI för arbetsminne underlätta ytterligare tolkning av resultaten. Tidigare studier har visat att cirkadiska bioritmer påverkar beteendeprestanda, och det finns individuella skillnader (Montplaisir, 1981; Lavie, 2001). Vi registrerade inte EEG-data samtidigt i detta experimentella, så påverkan av cirkadiska bioritmer på testresultaten kan inte helt uteslutas.
denna forskning visade att arbetsminneförmågan försämrades efter TSD och att denna skada inte var förknippad med stimulansinnehållet i arbetsminnet. Bristen på sömn minskade kvaliteten på informationen lagrad i minnet, vilket kan uppstå med degenerativ uppmärksamhetsprocess (Ratcliff och Van Dongen, 2018). Denna studie ger elektrofysiologiska bevis för att förstå mekanismen under det försämrade arbetsminnet efter sömnbrist. Det är nödvändigt att vara uppmärksam på de negativa effekterna av nedsatt arbetsminne som orsakas av sömnbrist och att undersöka effektiva ingrepp för sådan skada.
datatillgänglighet uttalande
de datamängder som genereras för denna studie är tillgängliga på begäran till motsvarande författare.
Etikuttalande
studierna som involverade mänskliga deltagare granskades och godkändes av det fjärde militära medicinska universitetet Beihang University. Patienterna / deltagarna gav sitt skriftliga informerade samtycke till att delta i denna studie.
Författarbidrag
Ys utformade experimenten. ZP producerade resultaten och skrev manuskriptet. CD och LZ analyserade och tolkade data. JT och YS utförde experimenten, bekantade data och garanterna för denna studie. YB, LZ och JT bidrog till att delta i datainsamling och granska litteraturen. Alla listade författare har läst och godkänt det slutliga manuskriptet.
finansiering
denna forskning stöddes av National Science Foundation of Winter Olympics Technology Plan of China under bidrag nr 2019yff0301600 och HJ20191A020135.
intressekonflikt
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
Anderson, C. och Platten, C. R. (2011). Sömnbrist sänker hämning och ökar impulsiviteten till negativa stimuli. Uppför dig. Hjärnan Res. 217: 466. doi: 10.1016 / j. bbr.2010.09.020
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Baddeley, A. (2000). Den episodiska bufferten: en ny komponent i arbetsminnet? Trender Cogn. Sci. 4:417. doi: 10.1016 / s1364-6613 (00) 01538-2
CrossRef fulltext/Google Scholar
Bartel, P., Offermeier, W., Smith, F. och Becker, P. (2004). Uppmärksamhet och arbetsminne i bosatta narkosläkare efter natttjänst: grupp-och individuella effekter. Occup. Miljö. Med. 61, 167–170. doi: 10.1136 / oem.2002.006197
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Casement, MD, Broussard, Jl, Mullington, JM och Press, Dz (2006). Sömnens bidrag till förbättringar i arbetsminneskanningshastighet: en studie av långvarig sömnbegränsning. Biol. Psychol. 72:212. doi: 10.1016/j.biopsycho.2005.11.002
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Chee, M. W. L. och Chuah, L. Y. M. (2008). Funktionell neuroimaging insikter i hur sömn och sömnbrist påverkar minne och kognition. Curr. Opin. Neurol. 21, 417–423. doi: 10.1097 / WCO.0b013e3283052cf7
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S. och Chee, M. W. L. (2005). Dissociation av kortikala regioner modulerade av både arbetsminnebelastning och sömnbrist och genom sömnbrist ensam. Neuroimage 25, 579-587. doi: 10.1016 / j. neuroimage.2004.11.029
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Chuah, YML, Venkatraman, V., Dinges, D. F. och Chee, M. W. L. (2006). Den neurala grunden för interindividuell variation i hämmande effektivitet efter sömnbrist. J. Neurosci. 26, 7156–7162. doi: 10.1523/jneurosci.0906-06. 2006
CrossRef fulltext/Google Scholar
Cote, K. A., Milner, ce, Osip, S. L., Baker, M. L. och Cuthbert, B. P. (2008). Fysiologisk upphetsning och uppmärksamhet under en vecka med kontinuerlig sömnbegränsning. Fysiol. Uppför dig. 95, 353–364. doi: 10.1016/j.physbeh.2008.06.016
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Cote, K. A., Mondloch, C. J., Sergeeva, V., Taylor, M. och Semplonius, T. (2014). Påverkan av total sömnbrist på beteendemässig neural bearbetning av känslomässigt uttrycksfulla ansikten. Exp. Hjärnan Res. 232, 1429-1442. doi: 10.1007 / s00221-013-3780-1
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Donchin, E. och Fabiani, M. (1991). Användningen av händelserelaterade hjärnpotentialer i studien av minne: är p300 ett mått på händelsens särskiljningsförmåga? Psychol. Facul. Offentlig. 7, 58–77. doi: 10.1111/sltb.12055
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Drummond, S. P., brun, G. G., Salamat, J. S. och Gillin, J. C. (2004). Ökande uppgiftssvårigheter underlättar det cerebrala kompensatoriska svaret på total sömnbrist. Sova 27, 445-451. doi: 10.1093 / sömn / 27.3.445
CrossRef fulltext / Google Scholar
Drummond, S. P. A. och Brown, G. G. (2001). Effekterna av total sömnbrist på cerebrala svar på kognitiv prestanda. Neuropsykofarmakologi 25, S68-S73. doi: 10.1016 / S0893-133x (01)00325-6
CrossRef fulltext / Google Scholar
Evans, km och Federmeier, KD (2007). Minnet som är rätt och minnet som är kvar: händelserelaterade potentialer avslöjar hemisfäriska asymmetrier vid kodning och lagring av verbal information. Neuropsykologi 45, 1777-1790. doi: 10.1016/j.neuropsykologi.2006.12.014
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M. och h Acibller, Y. (2007). Visuell P2-komponent är relaterad till theta-faslåsning. Neurosci. Lett. 426, 181–186. doi:10.1016 / j. neulet.2007.08.062
PubMed Abstract | CrossRef fulltext/Google Scholar
g Bikraldine, R., Desgranges, B., Foret, J. och Eustache, F. (2005). Förhållandena mellan minnessystem och sömnstadier. J. Sova Res. 14, 123-140. doi: 10.1111 / j. 1365-2869. 2005.00450.X
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Gosselin, A., Koninck, jd och Campbell, KB (2005). Total sömnbrist och nyhetsbehandling: konsekvenser för frontallobens funktion. Clin. Neurofysiol. 116, 211–222. doi: 10.1016/j.clinph.2004.07.033
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Harrison, Y. och Horne, J. (1998). Sömnförlust försämrar korta och nya språkuppgifter som har ett prefrontalt fokus. J. Sova Res. 7, 95-100. doi: 10.1046 / j. 1365-2869. 1998.00104.X
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J., och Meier, B. (2010). Den samtidiga giltigheten av N-back-uppgiften som ett arbetsminnesmått. Minne 18, 394-412. doi: 10.1080 / 09658211003702171
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N. och Chamari, K. (2013). Effekten av partiell sömnbrist på de dagliga variationerna av kognitiv prestanda hos utbildade ämnen. Proc. Soc. Uppför dig. Sci. 82, 392–396. doi: 10.1016/j.sbspro.2013.06.281
CrossRef fulltext / Google Scholar
Jennings, J. R., Monk, T. H. och Molen, M. W. V. D. (2003). Sömnbrist påverkar vissa men inte alla processer av tillsynsuppmärksamhet. Psychol. Sci. 14, 473–479. doi: 10.1111/1467-9280. 02456
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L. och Lei, Y. (2015). Recovery sleep reverserar nedsatt responshämning på grund av sömnbegränsning: bevis från en visuell händelserelaterad potentialstudie. PLoS One 10:e0142361. doi: 10.1371 / tidskrift.pone.0142361
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Jones, K. och Harrison, Y. (2001). Frontallobfunktion, sömnförlust och fragmenterad sömn. Sova Med. Rev. 5: 475. doi: 10.1053 / smrv.2001.0203
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Koslowsky, M. och Babkoff, H. (1992). Meta-analys av förhållandet mellan total sömnbrist och prestanda. Chronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A. och Hansenne, M. (2014). Alkoholmissbruk och erp-komponenter i go/no-go-uppgifter med alkoholrelaterade stimuli: påverkan av alkoholundvikande. Int. J. Psykofysiol. 94, 92–99. doi: 10.1016/j.ijpsycho.2014.08.001
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F. och Huster, R. J. (2019). Sömnbrist påverkar differentiellt delKomponenter av kognitiv kontroll. Sova 42: zsz016. doi: 10.1093 / sömn / zsz016
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Lavie, P. (2001). Sömn-vakna som en biologisk rytm. Annu. Pastor Psychol. 52, 277–303. doi: 10.1146 / annurev.psych.52.1.277
CrossRef fulltext / Google Scholar
Lei, Y., Shao, Y., Wang, L., Zhai, T. och Zou, F. (2015). Storskalig hjärnnätverkskoppling förutsäger totala sömnberövande effekter på kognitiv kapacitet. PLoS En 10: e0133959. doi: 10.1371 / tidskrift.pone.0133959
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Li, C., Huang, D., Qi, J., Chang, H. och Meng, Q. (2016). Kortvariga minnesunderskott korrelerar med hippocampal-talamiska funktionella anslutningsförändringar efter akut sömnbegränsning. Brain Imaging Beter Sig. 11, 954–963. doi: 10.1007 / s11682-016-9570-1
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Ma, N., Dinges, D. F., Basner, M. och Rao, H. (2014). Hur akut Total sömnförlust påverkar den närvarande hjärnan: en metaanalys av neuroimagingstudier. Sova 38, 233. doi: 10.5665 / sömn.4404
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Miyake, A. och Shah, P. (1999). Modeller av arbetsminne. Mekanismer för aktivt Underhåll och verkställande kontroll. Cambridge: Cambridge University Press.
Google Scholar
Mograss, ma, Guillem, F., Brazzini-Poisson, V. och Godbout, R. (2009). Effekterna av total sömnbrist på igenkänningsminnesprocesser: en studie av händelserelaterad potential. Neurobiol. Lära. Mem. 91, 343–352. doi: 10.1016 / j. nlm.2009.01.008
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Montplaisir, J. (1981). Depression och biologisk rytm: terapeutiska effekter av sömnbrist. L ’ Uni ubign m Ubidicale Du Kanada 110, 272-276. doi: 10.1620/tjem.133.481
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Morris, A. M., så, Y., Lee, K. A., Lash, A. A. och Becker, C. E. (1992). Den P300 händelse-relaterade potential. effekterna av sömnbrist. J. Ockupera. Med. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F. och Hobson, J. A. (2002). Den prefrontala cortex i sömn. Trender Cogn. Sci. 6, 475–481. doi: 10.1016 / S1364-6613 (02) 01992-7
CrossRef fulltext | Google Scholar
Ning, M., Dinges, D. F., Mathias, B. Och Hengyi, R. (2014). Hur akut Total sömnförlust påverkar den närvarande hjärnan: en metaanalys av neuroimagingstudier. Sova 38, 233-240. doi: 10.5665 / sömn.4404
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, A. R. och Bullmore, E. (2005). N-back arbetsminnesparadigm: en metaanalys av normativa funktionella neuroimagingstudier. Brum. Brain Mapp. 25, 46–59. doi: 10.1002 / hbm.20131
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S. och Kumar, S. (2010). Elektrofysiologiska korrelat av kognition förbättras med tupplur under sömnbrist. EUR. J. Appl. Fysiol. 108, 549–556. doi: 10.1007 / s00421-009-1222-3
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Ratcliff, R. och Van Dongen, H. (2018). Effekterna av sömnbrist på objekt och associativt igenkänningsminne. J. Exp. Psychol. 44, 193–208. doi: 10.1037 / xlm0000452
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Raymond, ca (1988). Skiftande arbete, sömncykler är på väg att bli en annan folkhälsofråga. JAMA J. Am. Med. Assoc. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef fulltext / Google Scholar
Saito, H., Yamazaki, H., Matsuoka, H., Matsumoto, K. och Sato, M. (2001). Visuell händelserelaterad potential vid mild demens av Alzheimers typ. Psykiatri Clin. Neurosci. 55, 365–371. doi: 10.1046 / j. 1440-1819. 2001.00876.X
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Schacht, A., Werheid, K., och Sommer, W. (2008). Bedömningen av ansiktsskönhet är snabb men inte obligatorisk. Cogn. Påverka. Uppför dig. Neurosci. 8, 132–142. doi: 10.3758 / cabn.8.2.132
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Scott, J. P. R., Mcnaughton, L. R. och Polman, R. C. J. (2006). Effekter av sömnbrist och motion på kognitiv, motorisk prestanda och humör. Fysiol. Uppför dig. 87:408. doi: 10.1016/j.physbeh.2005.11.009
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Smallwood, J., Brown, K., Baird, B. Och Schooler, J. W. (2011). Samarbete mellan default mode network och frontal-parietal network i produktionen av ett internt tankesätt. Hjärnan Res. 1428, 60-70. doi: 10.1016/j.brainres.2011.03.072
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Smith, Me, Mcevoy, lk och Gevins, A. (2002). Effekten av måttlig sömnförlust på neurofysiologiska signaler under arbetsminneuppgiftsprestanda. Sova 25, 784-794. doi: 10.1093 / sömn / 25.7.56
CrossRef fulltext / Google Scholar
Thomas, M. (2003). Neural grund av vakenhet och kognitiva funktionsnedsättningar under sömnighet ii. effekter av 48 och 72 timmar av sömnbrist på att vakna mänsklig regional hjärnaktivitet. Thalam. Relat. Syst. 2, 199–229. doi: 10.1016 / S1472-9288(03) 00020-7
CrossRef fulltext | Google Scholar
Van Dongen, H. (2004). Den kumulativa kostnaden för ytterligare vakenhet: dosesponse effekter på neurobeteendefunktioner och sömnfysiologi från kronisk sömnbrist och total sömnbrist. Sova 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef fulltext / Google Scholar
Verweij, im, Romeijn, N., Smit, DJ och Giovanni, P. (2014). Sömnbrist leder till förlust av funktionell anslutning i frontala hjärnregioner. BMC Neurosci. 15:88. doi: 10.1186/1471-2202-15-88
PubMed Abstrakt | CrossRef fulltext/Google Scholar
Wen, X., Liu, Y., Yao, L. och Ding, M. (2013). Top-down reglering av standardläge aktivitet i rumslig visuell uppmärksamhet. J. Neurosci. 33, 6444–6453. doi: 10.1523/JNEUROSCI.4939-12. 2013
CrossRef fulltext / Google Scholar
Wiggins, E., Malayka, M. och Kendra, G. (2018). 24-h sömnbrist försämrar tidig attentional modulering av neural bearbetning: en händelserelaterad hjärnpotentialstudie. Neurosci. Lett. 677, 32–36. doi:10.1016 / j. neulet.2018.04.022
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y. och Zhou, Q. (2019). Minskad information ersättning av arbetsminne efter sömnbrist: bevis från en händelserelaterad potentiell studie. Front. Neurosci. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef fulltext / Google Scholar
Zhang, Q., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z., et al. (2014). En visuell erp-studie av impulshämning efter en zaleplon-inducerad tupplur efter sömnbrist. PLoS en 9: e95653. doi: 10.1371 / tidskrift.pone.0095653
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Leave a Reply