frontiere în neuroștiințe
Introducere
odată cu progresul societății și schimbările în ritmul de lucru, un număr tot mai mare de oameni suferă de privarea de somn. Privarea de somn nu numai că dăunează sănătății fizice și mentale a individului, dar afectează grav și performanța la locul de muncă, provocând erori de muncă și chiar accidente. Prin urmare, înțelegerea mecanismului privării de somn care afectează funcția cognitivă este de mare importanță pentru prevenirea eficientă a efectelor privării de somn.
studiile anterioare au arătat că privarea de somn poate provoca o serie de schimbări în starea de spirit a unui individ, capacitatea cognitivă, performanța la locul de muncă și funcția imună (Choo și colab., 2005). Lipsa somnului perturbă circulația corpului și afectează abilitățile cognitive și emoționale ale indivizilor (Raymond, 1988). Mai multe studii au arătat că privarea de somn afectează inhibarea răspunsului (Harrison și Horne, 1998; Muzur și colab., 2002; Jennings și colab., 2003). De exemplu, după 36 de ore de privare de somn, capacitatea individului de a suprima stimulii negativi a scăzut (Chuah și colab., 2006). Studiile de neuroimagistică au sugerat că privarea de somn reduce nivelul scăzut al capacității de procesare vizuală a unui individ (Anderson și Platten, 2011; Ning și colab., 2014). În plus, privarea de somn afectează hipocampul și ar putea afecta memoria prin distrugerea plasticității sinaptice (Cote și colab., 2014). Thomas (2003) a indicat faptul că lipsa somnului a redus fluxul sanguin cerebral și rata metabolică în talamus, cortexul prefrontal și cortexul parietal (G Ecacraldine și colab., 2005). Jarraya și colegii săi au descoperit că privarea parțială de somn a afectat semnificativ funcțiile neuropsihologice, cum ar fi memoria verbală instantanee, atenția și vigilența (Thomas, 2003). Mai mult, unele studii au arătat că efectele cumulative ale privării parțiale de somn ar putea afecta grav funcția și comportamentul cognitiv (Van Dongen, 2004; Scott și colab., 2006; Jarraya și colab., 2013).
memoria de lucru este un sistem folosit pentru stocarea și procesarea informațiilor și care este o funcție cognitivă cu capacitate limitată (Bartel și colab., 2004). Mai mult, informațiile stocate în sistemul de memorie de lucru pot fi schimbate din memoria pe termen scurt în memoria pe termen lung prin repovestire și alte metode de memorie. Memoria de lucru este tranziția între sistemele de memorie pe termen scurt și pe termen lung, care este foarte esențială în procesarea mesajelor umane (Miyake și Shah, 1999). Oferă un spațiu de stocare temporar și resursele necesare pentru procesarea informațiilor, cum ar fi înțelegerea vocală, raționamentul și învățarea. Privarea de somn s-a dovedit a afecta mai întâi memoria de lucru.
studiile anterioare au folosit paradigma memoriei de lucru N-back la participanții care au suferit privarea de somn și au constatat că lipsa somnului induce o scădere a activității metabolice în rețeaua regională a creierului, care se efectuează în principal procesarea informațiilor și inhibarea reacției (Baddeley, 2000; Zhang și colab., 2019). Memoria de lucru afectată după privarea de somn este legată de activarea rețelei implicite în SARCINI (Chee și Chuah, 2008), care poate fi legată de rolul important al talamusului în vigilența corticală. De exemplu, privarea de somn a crescut legătura dintre hipocamp, talamus și rețeaua implicită, care a fost adesea însoțită de somnolență subiectivă mai mare și performanțe mai slabe ale memoriei de lucru (Lei și colab., 2015; Li și colab., 2016). Studii privind privarea de somn identi?că latența crescută și amplitudinea redusă a componentei P3 au fost asociate cu sobrietatea prelungită (Morris și colab., 1992; Jones și Harrison, 2001; Panjwani și colab., 2010). Scăderea undei P3 ar putea reflecta o scădere a atenției participanților și o reducere a discernământului stimulilor țintă (Koslowsky și Babkoff, 1992).cu toate acestea, puține studii au furnizat dovezi electrofiziologice pentru afectarea memoriei de lucru după privarea de somn. Sarcina N-back este considerată o metodă comună de evaluare a memoriei de lucru (Owen și colab., 2005; Jaeggi și colab., 2010). Zhang și colab. a proiectat o sarcină de memorie de lucru cu două spate pentru a explora supleantul de mesaje scăzut al memoriei de lucru în timpul privării de somn, dar puține studii au folosit diferite tipuri de sarcini de memorie de lucru într-un singur experiment. În studiul de față, am proiectat diferite tipuri de sarcini de memorie de lucru (memoria de lucru a pronunției, memoria de lucru spațială și memoria de lucru a obiectelor) pentru a explora afectarea funcției cognitive de către TSD și a înregistrat datele EEG ale participanților la 2 puncte de timp (baseline și 36 h-TSD). Toate sarcinile au adoptat o paradigmă cu 2 spate. Acest studiu a evaluat modificările undei N2-P3 legate de memoria de lucru în timpul TSD și a analizat caracteristicile temporale ale efectelor privării de somn asupra memoriei de lucru. Descoperirile noastre oferă dovezi experimentale pentru efectele privării de somn asupra funcției cognitive.
materiale și metode
participanți
șaisprezece tineri, sănătoși, studenți de sex masculin dreptaci au participat la acest studiu. Am recrutat participanți prin publicitate în campus. Toți participanții au avut obiceiuri bune de somn (PSQI<5). Toți participanții au avut vârsta cuprinsă între 21 și 28 de ani, cu o vârstă medie de 23 de ani și niciunul dintre participanți nu a avut nicio boală mentală sau fizică. Toți participanții au avut viziune normală sau viziune corectată peste 1,0 și scoruri de inteligență >110 la testul Raven. Înainte de experiment, experimentatorul a explicat procedura și punctele de atenție participanților pentru a se asigura că sunt familiarizați cu metoda și procedura. În cele 2 săptămâni anterioare experimentului, participanții au dormit în mod regulat timp de 7-9 ore pe zi, fără să fumeze, să bea cafea, să bea alcool sau să consume orice medicament timp de 2 zile înainte de experiment. Înainte de experiment, toți participanții au oferit consimțământul scris în cunoștință de cauză. Schema experimentală a fost aprobată de Comitetul de etică al celei de-a patra Universități Medicale Militare și de Universitatea Beihang.
design Experimental
trei tipuri de sarcini de memorie de lucru au fost prezentate tuturor participanților. Au fost sarcină de memorie de lucru cu două spate (a se vedea Figura 1), sarcină de memorie de lucru spațială cu două spate (a se vedea Figura 2) și sarcină de memorie de lucru cu două spate (a se vedea Figura 3). Materialele stimulante ale sarcinilor au fost 15 litere englezești insensibile la majuscule, excluzându-le pe cele cu litere similare, cum ar fi L/l, M/m; pătrate negre mici; și, respectiv, 12 figuri geometrice. Toate materialele au fost prezentate în culoarea neagră pe un fundal alb, cu un unghi vizual aproximativ de 1,5 XCT 1,5 XCT (lățime: 2,0 cm, înălțime: 2,0 cm) subtending. 122 trasee au fost cuprinse în fiecare sarcină și, în fiecare traseu, stimulul țintă a fost prezentat pentru 400 ms două trasee după prezentarea stimulului obiectiv, cu 1.600 ms stimul debut asincronie timp (SOA) care a fost marcat de un alb „+.Participanții au fost rugați să facă clic pe butonul stâng al mouse-ului atunci când ținta și stimulul obiectiv au fost aceleași („potrivire”), în timp ce faceți clic pe butonul din dreapta al mouse-ului atunci când nu au fost („nepotrivire”). Condiția de potrivire sau nu a fost prezentată într-o ordine pseudorandom cu un raport 1:1.
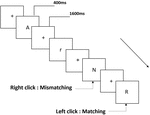
Figura 1. Diagrama schematică a sarcinii de memorie de lucru a pronunției.
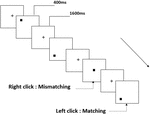
Figura 2. Diagrama schematică a sarcinii memoriei de lucru spațiale.
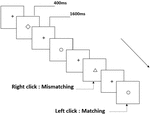
Figura 3. Diagrama schematică a sarcinii de memorie de lucru obiect.
proceduri experimentale
înainte de experiment, participanții au fost instruiți cu privire la sarcina experimentală. Ei au fost informați să practice cele trei tipuri de sarcini de memorie de lucru până când a fost atinsă o rată de precizie de 90%. Participanții au vizitat Laboratorul cu o zi înainte de experiment și au dormit în laborator în acea noapte. Cei doi participanți parteneri au efectuat experimentele în același timp. Trei tipuri de sarcini de memorie de lucru au fost efectuate la 7:30 până la 8:30 dimineața următoare, cu înregistrare simultană a electroencefalogramei (EEG) (linia de bază). A doua înregistrare EEG (36 h-TSD) a fost efectuată după o perioadă de 36 de ore în care participanților nu li sa permis să doarmă. Pe tot parcursul experimentului, inhibiția Centrală și medicamentele stimulante au fost interzise. Participanții au fost însoțiți, observați și reamintiți de personalul medical pentru a-i menține treji pe tot parcursul sesiunii TSD.
înregistrări EEG
un EEG continuu al scalpului a fost înregistrat folosind capace de electrozi plasate în 64 de locații folosind sistemul 10-20 cu un amplificator SynAmps2. Mastoidele bilaterale (A1 și A2) au fost folosite pentru referință, iar fruntea a fost folosită ca sol. EEG-urile au fost înregistrate la 1.000 Hz, iar impedanța tuturor canalelor a fost menținută sub 5 k. Patru electrozi suplimentari au fost plasați deasupra și dedesubtul ochilor drepți și stângi pentru a înregistra o electrooculogramă verticală și orizontală bipolară.
analiza datelor experimentelor comportamentale
Din cauza erorilor tehnice, două cazuri au fost șterse, în timp ce alte 14 cazuri au fost incluse în următoarea analiză statistică. Datele comportamentale au inclus timpul mediu de reacție, rata corectă și numărul corect pe unitate de timp. Datele comportamentale în starea inițială și 36 de stări h-TSD au fost înregistrate pentru analiză. Analizele au fost efectuate de IBM SPSS (V22.2), în cazul în care au fost lansate măsurile repetate de analiză a varianței (ANOVA) metoda cu efect de seră-Geisser Bonferroni analiza post-hoc. Rezultatele statistice au fost prezentate ca medie și deviație standard (DS).
analiza datelor EEG
programul Scan 4.3 a fost utilizat pentru a analiza datele EEG, unde artefactele EEG ale mișcării ochilor au fost corectate prin metoda reducerii artefactelor oculare. Epocile cuprinse între -100 și 800 ms din datele EEG continue au fost extrase și filtrate de un filtru de bandă de la 0,5 la 30 Hz cu panta de frecvență de 24 dB/oct. Studiile în care tensiunea a depășit valoarea de 100 la sută la sută au fost respinse, iar valoarea inițială a fost corectată la o amplitudine medie de 100 ms. componentele EEG au fost mediate și calculate doar cu răspunsurile corectate. Componentele ERP P2 (100-250 ms), N2 (150-350 ms) și P3 (250-450 ms) ale studiilor de stimulare au fost identificate și cuantificate. Amplitudinile și latențele maxime medii ale componentelor N2 și P3 au fost calculate separat la F3, Fz, F4, C3, Cz, C4, P3, Pz și P4, iar componenta P2 a fost calculată la F3, Fz, F4, C3, Cz și C4 (Casement și colab., 2006; Verweij și colab., 2014).
măsuri repetate ANOVA a fost utilizată pentru toate rezultatele ERP. Efectele principale și interacțiunile dintre stările de somn (baseline și 36 h-TSD), sarcinile (memoria de lucru a pronunției, memoria de lucru spațială și memoria de lucru a obiectelor), regiunile (frontal, central și parietal; componenta P2 a fost analizată doar pe regiunile frontale și centrale) și siturile (stânga, mijloc și dreapta) au fost analizate statistic folosind măsuri repetate ANOVA, care au inclus corecții de seră-Geisser pentru teste non-sfericitate și Bonferroni post-hoc.
rezultate
performanță comportamentală
rezultatele experimentelor comportamentale sunt prezentate în tabelul 1. Timpul mediu de reacție a fost mai lung în starea 36 h-TSD decât la momentul inițial, cu o tendință de creștere, dar fără diferențe semnificative. ANOVA a arătat că rata corectă a sarcinii a fost semnificativ diferită între linia de bază și 36 h-TSD . Numărul corect pe unitate de timp a arătat un efect principal semnificativ al timpului în timpul 36 h-TSD .

Tabelul 1. Date de performanță (SD medie de la sută la sută) pentru sarcina cu 2 spate la momentul inițial și după 36 h-TSD.
amplitudine
comparativ cu valoarea inițială, s-a observat o scădere semnificativă a amplitudinii P3 și s-a observat o creștere semnificativă a amplitudinii P2 după TSD. Deși amplitudinea N2 a scăzut după 36 de ore de TSD, diferența nu a atins semnificația statistică (Tabelul 2).

Tabelul 2. Grand-amplitudinea medie de vârf a componentelor P2, N2 și P3 în starea de Răspuns corectă pe mai multe site-uri de electrozi la momentul inițial și după 36 h-TSD.
principalele efecte semnificative ale regiunilor și siturilor asupra amplitudinii P2 au fost găsite în condiția TSD. În timpul TSD, amplitudinea maximă a P2 a apărut în regiunea frontală (Figura 4). În plus, diferențele în amplitudinile P2 în diferite regiuni (frontal vs.central) au fost mai semnificative la electrozii bilaterali (stânga: P = 0,001; dreapta: P = 0,000) decât la electrozii din mijloc (Figura 4). Un efect principal semnificativ al regiunii și sitului asupra N2 a arătat că amplitudinea N2 a fost mai negativă în regiunea frontală decât în regiunea centrală (P = 0,008, Figura 4) și a fost mai mică în dreapta decât în partea stângă (P = 0,011, figurile 5A2,B2). A fost observat un efect principal al site-ului asupra amplitudinii P3 . Amplitudinea P3 a fost mai pozitivă la mijloc decât pe partea stângă (P = 0,009, Figura 4). S-a observat un efect semnificativ de interacțiune între timp și regiune pentru amplitudinea P3 . În timpul TSD, reducerea amplitudinii P3 a fost mai semnificativă în regiunile frontale și centrale decât în regiunea parietală (P = 0,005; P = 0,003) (figura 5C3). Niciun alt efect principal sau efect de interacțiune nu a atins o semnificație statistică.
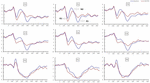
Figura 4. Amplitudinea ERP la momentul inițial și 36 h-TSD pentru starea de Răspuns corectă pentru sarcina de memorie de lucru. Canalele sunt ordonate de la stânga la dreapta și de sus în jos după cum urmează: F3, Fz, F4, C3, Cz, C4, P3, Pz și P4. Comparativ cu valoarea inițială, latențele componentelor N2-P3 au fost prelungite, iar amplitudinile N2-P3 au scăzut după 36 h-TSD.
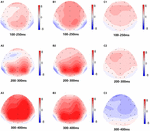
Figura 5. Harta topografică a răspunsului corect în sarcina memoriei de lucru în diferite condiții de somn (A1–C3). (A1) P2, 100-250 ms, la momentul inițial. (A2) N2, 200-300 ms, la momentul inițial. (A3) P3, 300-400 ms, la momentul inițial. (B1) P2, 100-250 ms, la 36 h-TSD. (B2) N2, 200-300 ms, pentru 36 h-TSD. (B3) P3, 300-400 ms, pentru 36 h-TSD. (C1) P2, 100-250 ms, 36 h-TSD cu valoarea inițială scăzută. (C2) N2, 200-300 ms, 36 h-TSD cu valoarea inițială scăzută. (C3) P3, 300-400 ms, 36 h-TSD cu valoarea inițială scăzută.
latență
latențele N2 și P2 au fost semnificativ prelungite după TSD. Deși latența P3 a fost prelungită după 36 de ore de TSD, diferența nu a atins semnificația statistică (Tabelul 3).

Tabelul 3. Grand-latența medie de vârf a componentelor P2, N2 și P3 în starea de Răspuns corectă pe mai multe site-uri de electrozi la momentul inițial și după 36 h-TSD.
efectul principal semnificativ al regiunii asupra N2 și P3 a arătat că latența componentelor N2-P3 a fost mai scurtă în regiunea parietală decât în regiunea frontală (P = 0,002; P = 0,000) și regiunea centrală (P = 0,000; P = 0,000) (Figura 4). Latența undei P3 a fost semnificativ mai lungă pe partea stângă decât pe partea dreaptă (Figura 4).
niciun alt efect principal sau efect de interacțiune nu a atins o semnificație statistică.
amplitudinile și latențele N2, P2 și P3 care au fost obținute la cele nouă locuri de electrozi sunt prezentate în Figura 4. Harta topografică a răspunsului corect în sarcina memoriei de lucru în diferite condiții de somn (baseline, 36 h-TSD și diferența dintre cele două condiții) este prezentată în Figura 5.
discuție
în acest studiu, am raportat influențele privării de somn de 36 de ore asupra memoriei de lucru, combinând datele comportamentale în două stări de somn (baseline și 36 h-TSD) cu înregistrări EEG contemporane. Analiza rezultatelor a indicat că modificările datelor comportamentale în conformitate cu memoria de lucru afectată după 36 h TSD: o creștere a timpului mediu de reacție al sarcinilor cognitive și o scădere a preciziei.
privarea de somn a afectat controlul individual al resurselor atenționale. Deși indivizii au încercat să mențină starea de veghe și performanța muncii, inclusiv timpul de reacție și rata corectă, în timpul privării de somn, capacitatea de procesare a informațiilor din memoria lor de lucru a fost încă afectată din cauza scăderii vitezei de procesare a informațiilor (Casement și colab., 2006; Wiggins și colab., 2018). În acest studiu, undele N2 și P3 legate de memoria de lucru au fost măsurate pentru a arăta o creștere a latenței și o scădere a amplitudinii după privarea de somn comparativ cu citirile de bază. Studiile au demonstrat că privarea de somn duce la o scădere continuă a atenției, iar fenomenul scăderii amplitudinii P3 indică faptul că controlul cogniției de sus în jos al indivizilor se prăbușește treptat. Privarea de somn are un efect mai advers asupra funcțiilor cognitive, în special a celor care depind de mental sau cogniții (Kusztor și colab., 2019).
componenta P3 reflectă desfășurarea resurselor de atenție, iar latența P3 este văzută pe scară largă ca fereastra de timp pentru clasificarea și evaluarea stimulilor. Scăderea amplitudinii undei P3 a confirmat, de asemenea, că luarea deciziilor în răspunsul de potrivire după TSD a fost deteriorată într-o anumită măsură (Gosselin și colab., 2005). Studiile au sugerat că privarea de somn poate afecta stadiul de procesare a informațiilor din memoria de lucru. În acest studiu, indicatorii de performanță au susținut, de asemenea, concluzia că timpul de răspuns la stimulul țintă a fost crescut și că latența undei P3 a fost prelungită (Cote și colab., 2008). S-a speculat că efectul privării de somn asupra componentelor P3 ar putea avea loc și din cauza eșecului de a răspunde la modificarea informațiilor, ceea ce este în concordanță cu concluziile anterioare conform cărora componentele P3 sunt legate de actualizarea conținutului memoriei de lucru (Donchin și Fabiani, 1991).
studiile anterioare au considerat componenta N2 ca un indice electrofiziologic care reflectă capacitatea individului de a suprima răspunsul (Kreusch și colab., 2014). După privarea de somn, latența prelungită a componentei NoGo-N2 indică faptul că capacitatea individului de a suprima răspunsul este afectată (Jin și colab., 2015). Amplitudinea scăzută și latența prelungită a componentei N2 legate de memoria de lucru a pronunției după privarea de somn arată că privarea de somn afectează procesarea informațiilor din memoria de lucru a pronunției (Zhang și colab., 2019). Componenta N2 este în general considerată a reflecta atenția selectivă a creierului și procesarea stimulilor sau semnalelor emoționale (Schacht și colab., 2008) și este o componentă endogenă legată de starea mentală, atenția și gradul de atenție al unui individ. În acest studiu, am constatat că latența componentei N2 a fost semnificativ prelungită, dar amplitudinea a arătat doar o tendință descendentă. Conform studiilor anterioare, latența prelungită a N2 a reflectat o creștere a timpului de răspuns după restricția de somn (Zhang și colab., 2014). Cu toate acestea, constatarea că amplitudinea N2 nu a fost modificată semnificativ s-ar fi putut datora răspunsurilor compensatorii cerebrale (Drummond și Brown, 2001). În cazul resurselor cognitive limitate, a existat un mecanism de compensare pentru a restabili funcția cognitivă afectată (Jin și colab., 2015).conform topografiei scalpului, modificările componentelor N2-P3 legate de privarea de somn sunt mai evidente în zona frontală. Controlul Frontoparietal (FPC) joacă un rol important în controlul cognitiv. Studiile au arătat că FPC poate ocoli controlul cognitiv de sus în jos, permițând indivizilor să se concentreze asupra informațiilor legate de țintă în timp ce suprimă informațiile care nu sunt legate de țintă (Smallwood și colab., 2011; Wen și colab., 2013). FPC este important pentru păstrarea informațiilor și prelucrarea informațiilor în memoria de lucru, iar gradul de activare a FPC după privarea de somn a fost redus în comparație cu un grup normal de somn (Ma și colab., 2014). Deși rezultatele EEG nu au reflectat în detaliu modificările din anumite regiuni ale creierului, au reflectat intuitiv efectul TSD asupra reținerii și procesării informațiilor din memoria de lucru.
deși procesul cognitiv exact care stă la baza componentei P2 este încă dezbătut pe scară largă, ca o definiție largă, componenta P2 reflectă procesul de atenție și procesare vizuală și este în general considerată a fi legată de atenția selectivă și memoria de lucru, reflectând judecata timpurie a procesului perceptiv (Saito și colab., 2001). În acest studiu, am constatat o creștere semnificativă a amplitudinii undei P2 după privarea de somn. Studiile au raportat că undele P2 care ar putea face parte din sistemul de potrivire cognitivă timpurie pentru procesarea mesajelor și pot compara intrările senzoriale cu memoria stocată (Freunberger și colab., 2007) sunt sensibile la modificările atenției misiunii și ale cerințelor memoriei de lucru (Smith și colab., 2002). Compensarea funcțională este una dintre funcțiile unice ale creierului uman și un factor important pentru menținerea funcției cognitive. Activarea excesivă a cortexului prefrontal dorsolateral (DLPFC) după privarea de somn indică faptul că, pe măsură ce resursele creierului scad, DLPFC pare să aibă o funcție compensatorie (Drummond și colab., 2004; Choo și colab., 2005). Prin urmare, speculăm că creșterea semnificativă a amplitudinii P2 observată în acest studiu se poate datora compensării funcționale în care indivizii par să mențină funcția cognitivă normală după privarea de somn. Deși un număr mare de studii au folosit tehnologia ERP pentru a explora efectul privării de somn asupra funcțiilor cognitive, componentele timpurii precum N1 și P2 nu au fost studiate sistematic, iar rezultatele sunt inconsistente (Evans și Federmeier, 2007; Wiggins și colab., 2018; Zhang și colab., 2019). Există puține cercetări care explorează schimbarea componentei P2 în timpul privării de somn (Mograss și colab., 2009). Prin urmare, efectele privării de somn asupra componentelor timpurii ale ERP, cum ar fi P2, trebuie încă studiate și explorate în continuare.
în acest experiment, am folosit modelul 2-back pentru a proiecta sarcini de pronunție, spațiu și memorie de lucru a obiectelor și am examinat afectarea memoriei de lucru după 36 de ore de TSD. Comparativ cu studiile anterioare care s-au concentrat doar pe efectele privării de somn asupra unui anumit tip de informații, cum ar fi memoria de lucru a pronunției sau funcția cognitivă specifică, cum ar fi inhibarea răspunsului, am luat în considerare conținutul modelului de memorie de lucru și am analizat în mod cuprinzător efectele privării de somn asupra memoriei de lucru.
cu toate acestea, studiul are unele limitări. În primul rând, am folosit doar sarcina cu 2 spate și nu am reușit să comparăm performanța participanților la sarcinile de memorie de lucru cu diferite dificultăți. Prin urmare, există limitări în explicarea și deducerea modificărilor volumului de muncă. În al doilea rând, numai voluntarii de sex masculin au fost utilizați în studiu, iar concluziile trebuie evaluate atunci când le extindem la voluntarii de sex feminin. Datorită numărului limitat de participanți, am constatat doar că amplitudinea undei N2 a avut o tendință descendentă și că latența undei P3 a avut o tendință prelungită. Rezultate stabile pot fi obținute după creșterea numărului de participanți. În al treilea rând, combinarea procedurii noastre cu fMRI pentru memoria de lucru poate facilita interpretarea ulterioară a rezultatelor. Studiile anterioare au arătat că bioritmurile circadiene afectează performanța comportamentală și există diferențe individuale (Montplaisir, 1981; Lavie, 2001). Nu am înregistrat datele EEG în același moment în acest experimental, astfel încât influența bioritmurilor circadiene asupra rezultatelor testului nu poate fi exclusă complet.
această cercetare a arătat că capacitatea memoriei de lucru a fost afectată după TSD și că această deteriorare nu a fost asociată cu conținutul stimulului memoriei de lucru. Lipsa somnului a redus calitatea informațiilor stocate în memorie, care ar putea apărea odată cu procesul degenerativ de atenție (Ratcliff și Van Dongen, 2018). Acest studiu oferă dovezi electrofiziologice pentru înțelegerea mecanismului sub memoria de lucru afectată după privarea de somn. Este necesar să se acorde atenție efectelor adverse ale afectării memoriei de lucru cauzate de privarea de somn și să se exploreze intervenții eficiente pentru astfel de daune.
Declarație privind disponibilitatea datelor
seturile de date generate pentru acest studiu sunt disponibile la cerere autorului corespondent.
declarație de etică
studiile care au implicat participanți umani au fost revizuite și aprobate de a patra universitate medicală militară Universitatea Beihang. Pacienții / participanții și-au dat consimțământul scris în cunoștință de cauză pentru a participa la acest studiu.
contribuții autor
YS proiectat experimentele. ZP a produs rezultatele și a scris manuscrisul. CD și LZ au analizat și interpretat datele. JT și YS au efectuat experimentele, au familiarizat datele și garanții acestui studiu. YB, LZ și JT au contribuit la participarea la colectarea datelor și la revizuirea literaturii. Toți autorii enumerați au citit și aprobat manuscrisul final.
finanțare
această cercetare a fost susținută de Fundația Națională de știință a planului tehnologic al Jocurilor Olimpice de iarnă din China în cadrul grantului nr.2019yff0301600 și HJ20191A020135.
Conflict de interese
autorii declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Anderson, C., și Platten, C. R. (2011). Privarea de somn scade inhibarea și sporește impulsivitatea la stimulii negativi. Comportă-te. Brain Res. 217: 466. doi: 10.1016 / j. bbr.2010.09.020
PubMed Abstract | CrossRef Full Text/Google Scholar
Baddeley, A. (2000). Tamponul episodic: o nouă componentă a memoriei de lucru? Tendințe Cogn. Sci. 4:417. doi: 10.1016/s1364-6613(00)01538-2
CrossRef Full Text/Google Scholar
Bartel, P., Offermeier, W., Smith, F., și Becker, P. (2004). Atenție și memorie de lucru la anesteziștii rezidenți după datoria de noapte: efecte de grup și individuale. Occup. Environ. Med. 61, 167–170. doi: 10.1136/oem.2002.006197
PubMed Abstract | CrossRef Full Text/Google Scholar
Casement, MD, Broussard, Jl, Mullington, JM, and Press, Dz (2006). Contribuția somnului la îmbunătățirea vitezei de scanare a memoriei de lucru: un studiu al restricției prelungite a somnului. Biol. Psihol. 72:212. doi: 10.1016 / j. biopsiho.2005.11.002
PubMed Abstract | CrossRef Full Text/Google Scholar
Chee, M. W. L., și Chuah, L. Y. M. (2008). Informații neuroimagistice funcționale despre modul în care somnul și privarea de somn afectează memoria și cunoașterea. Curr. Opin. Neurol. 21, 417–423. doi: 10.1097 / OMV.0b013e3283052cf7
PubMed Abstract | CrossRef Full Text/Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S. și Chee, M. W. L. (2005). Disocierea regiunilor corticale modulate atât de sarcina memoriei de lucru, cât și de privarea de somn și numai de privarea de somn. Neuroimagine 25, 579-587. doi: 10.1016 / j. neuroimage.2004.11.029
PubMed Abstract | CrossRef Full Text/Google Scholar
Chuah, Y. M. L., Venkatraman, V., Dinges, D. F. și Chee, M. W. L. (2006). Baza neuronală a variabilității interindividuale în eficiența inhibitoare după privarea de somn. J. Neurosci. 26, 7156–7162. doi: 10.1523/jneurosci.0906-06.2006
CrossRef Full Text/Google Scholar
Cote, K. A., Milner, C. E., Osip, S. L., Baker, M. L. și Cuthbert, B. P. (2008). Excitare fiziologică și atenție în timpul unei săptămâni de restricție continuă a somnului. Physiol. Comportă-te. 95, 353–364. doi: 10.1016 / j. physbeh.2008.06.016
PubMed Abstract | CrossRef Full Text/Google Scholar
Cote, K. A., Mondloch, C. J., Sergeeva, V., Taylor, M. și Semplonius, T. (2014). Impactul privării totale de somn asupra procesării neuronale comportamentale a fețelor expresive emoțional. Exp. Brain Res. 232, 1429-1442. doi: 10.1007 / s00221-013-3780-1
PubMed Abstract | CrossRef Full Text/Google Scholar
Donchin, E. și Fabiani, M. (1991). Utilizarea potențialelor cerebrale legate de evenimente în studiul memoriei: este p300 o măsură a caracterului distinctiv al evenimentului? Psihol. Facul. Publice. 7, 58–77. doi: 10.1111 / sltb.12055
PubMed Abstract | CrossRef Full Text/Google Scholar
Drummond, S. P., Brown, G. G., Salamat, J. S. și Gillin, J. C. (2004). Creșterea dificultății sarcinii facilitează răspunsul compensator cerebral la privarea totală de somn. Somnul 27, 445-451. doi: 10.1093 / sleep/27.3.445
CrossRef Full Text / Google Scholar
Drummond, S. P. A., și Brown, G. G. (2001). Efectele privării totale de somn asupra răspunsurilor cerebrale la performanța cognitivă. Neuropsihofarmacologie 25, S68-S73. doi: 10.1016/S0893-133x(01)00325-6
CrossRef Full Text/Google Scholar
Evans, K. M., și Federmeier, K. D. (2007). Memoria care este dreapta și memoria care este stânga: potențialele legate de evenimente dezvăluie asimetrii emisferice în codificarea și reținerea informațiilor verbale. Neuropsihologia 45, 1777-1790. doi: 10.1016 / j. neuropsihologia.2006.12.014
PubMed Abstract | CrossRef Full Text/Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M., și H Unicller, Y. (2007). Componenta vizuală p2 este legată de blocarea fazei theta. Neuroști. Let. 426, 181–186. doi: 10.1016 / j. neulet.2007.08.062
PubMed Abstract | CrossRef Full Text/Google Scholar
g Unacraldine, R., Desgranges, B., Foret, J., și Eustache, F. (2005). Relațiile dintre sistemele de memorie și etapele de somn. J. Sleep Res. 14, 123-140. doi: 10.1111 / j. 1365-2869.2005. 00450.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Gosselin, A., Koninck, J. D. și Campbell, K. B. (2005). Privarea totală de somn și procesarea noutății: implicații pentru funcționarea lobului frontal. Clin. Neurofiziol. 116, 211–222. doi: 10.1016 / j. clinph.2004.07.033
PubMed Abstract | CrossRef Full Text/Google Scholar
Harrison, Y., și Horne, J. (1998). Pierderea somnului afectează sarcinile lingvistice scurte și noi, având un accent prefrontal. J. Sleep Res. 7, 95-100. doi: 10.1046 / j. 1365-2869.1998. 00104.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J., și Meier, B. (2010). Validitatea concurentă a sarcinii N-back ca măsură de memorie de lucru. Memoria 18, 394-412. doi: 10.1080 / 09658211003702171
PubMed Abstract | CrossRef Full Text/Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., și Chamari, K. (2013). Impactul privării parțiale de somn asupra variațiilor diurne ale performanței cognitive la subiecții instruiți. Proc. Soc. Comportă-te. Sci. 82, 392–396. doi: 10.1016 / j. sbspro.2013.06.281
CrossRef Full Text/Google Scholar
Jennings, J. R., Monk, T. H. și Molen, M. W. V. D. (2003). Privarea de somn influențează unele, dar nu toate procesele de atenție de supraveghere. Psihol. Sci. 14, 473–479. doi: 10.1111/1467-9280.02456
PubMed Abstract | CrossRef Full Text/Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L. și Lei, Y. (2015). Somnul de recuperare inversează inhibarea răspunsului afectat din cauza restricției de somn: dovezi dintr-un studiu de potențiale legate de evenimente vizuale. PLoS One 10:e0142361. doi: 10.1371/jurnal.pone.0142361
PubMed Abstract | CrossRef Full Text/Google Scholar
Jones, K. și Harrison, Y. (2001). Funcția lobului Frontal, pierderea somnului și somnul fragmentat. Somn Med. Apocalipsa 5:475. doi: 10.1053 / smrv.2001.0203
PubMed Abstract | CrossRef Full Text/Google Scholar
Koslowsky, M. și Babkoff, H. (1992). Meta-analiza relației dintre privarea totală de somn și performanță. Chronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
PubMed Abstract | CrossRef Full Text/Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A. și Hansenne, M. (2014). Abuzul de alcool și componentele erp în sarcinile go / no-go folosind stimuli legați de alcool: impactul evitării alcoolului. Int. J. Psihofiziol. 94, 92–99. doi: 10.1016 / j. ijpsiho.2014.08.001
PubMed Abstract | CrossRef Full Text/Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F. și Huster, R. J. (2019). Privarea de somn afectează diferențiat subcomponentele controlului cognitiv. Somn 42: zsz016. doi: 10.1093 / sleep/zsz016
PubMed Abstract | CrossRef Full Text/Google Scholar
Lavie, P. (2001). Somn-trezire ca ritm biologic. Anu. Rev. Psychol. 52, 277–303. doi: 10.1146/annurev.psych.52.1.277
CrossRef Full Text/Google Scholar
Lei, Y., Shao, Y., Wang, L., Zhai, T. și Zou, F. (2015). Cuplarea rețelei cerebrale la scară largă prezice efectele totale ale privării de somn asupra capacității cognitive. PLoS Unul 10: e0133959. doi: 10.1371/jurnal.pone.0133959
PubMed Abstract | CrossRef Full Text/Google Scholar
Li, C., Huang, D., Qi, J., Chang, H., și Meng, Q. (2016). Deficitele de memorie pe termen scurt se corelează cu modificările conectivității funcționale hipocampal-talamice după restricția acută a somnului. Imagistica Creierului Se Comportă. 11, 954–963. doi: 10.1007 / s11682-016-9570-1
PubMed Abstract | CrossRef Full Text/Google Scholar
Ma, N., Dinges, D. F., Basner, M. și Rao, H. (2014). Cât de acută pierderea totală a somnului afectează creierul participant: o meta-analiză a studiilor de neuroimagistică. Somnul 38, 233. doi: 10.5665/somn.4404
PubMed Abstract | CrossRef Full Text/Google Scholar
Miyake, A. și Shah, P. (1999). Modele de memorie de lucru. Mecanisme de întreținere activă și Control executiv. Cambridge: Cambridge University Press.
Google Scholar
Mograss, M. A., Guillem, F., Brazzini-Poisson, V., și Godbout, R. (2009). Efectele privării totale de somn asupra proceselor de memorie de recunoaștere: un studiu al potențialului legat de evenimente. Neurobiol. Învață. Mem. 91, 343–352. doi: 10.1016 / j. nlm.2009.01.008
PubMed Abstract | CrossRef Full Text/Google Scholar
Montplaisir, J. (1981). Depresia și ritmul biologic: efectele terapeutice ale privării de somn. L ‘ uniqciftnmcifdicale du Canada 110, 272-276. doi: 10.1620 / tjem.133.481
PubMed Abstract | CrossRef Full Text | Google Scholar
Morris, A. M., deci, Y., Lee, K. A., Lash, A. A. și Becker, C. E. (1992). Potențialul legat de evenimente p300. efectele privării de somn. J. Occupat. Med. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F. și Hobson, J. A. (2002). Cortexul prefrontal în somn. Tendințe Cogn. Sci. 6, 475–481. doi: 10.1016/S1364-6613(02)01992-7
CrossRef Full Text/Google Scholar
Ning, M., Dinges, D. F., Mathias, B. și Hengyi, R. (2014). Cât de acută pierderea totală a somnului afectează creierul participant: o meta-analiză a studiilor de neuroimagistică. Somn 38, 233-240. doi: 10.5665/somn.4404
PubMed Abstract | CrossRef Full Text/Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, A. R. și Bullmore, E. (2005). Paradigma memoriei de lucru N-back: o meta-analiză a studiilor neuroimagistice funcționale normative. Zumzet. Brain Mapp. 25, 46–59. doi: 10.1002 / hbm.20131
PubMed Abstract | CrossRef Full Text/Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S. și Kumar, S. (2010). Corelațiile electrofiziologice ale cunoașterii se îmbunătățesc cu somnul în timpul privării de somn. Eur. J. Appl. Physiol. 108, 549–556. doi: 10.1007 / s00421-009-1222-3
PubMed Abstract | CrossRef Full Text/Google Scholar
Ratcliff, R. și Van Dongen, H. (2018). Efectele privării de somn asupra memoriei de recunoaștere a elementelor și asociative. J. Exp. Psihol. 44, 193–208. doi: 10.1037/xlm0000452
PubMed Abstract | CrossRef Full Text/Google Scholar
Raymond, C. A. (1988). Schimbarea muncii, ciclurile de somn sunt pe cale să devină o altă problemă de sănătate publică. JAMA J. Am. Med. Conf. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef Full Text/Google Scholar
Saito, H., Yamazaki, H., Matsuoka, H., Matsumoto, K. și Sato, M. (2001). Potențial vizual legat de evenimente în demența ușoară de tip alzheimer. Psihiatrie Clin. Neuroști. 55, 365–371. doi: 10.1046 / j. 1440-1819.2001. 00876.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Schacht, A., Werheid, K., și Sommer, W. (2008). Evaluarea frumuseții faciale este rapidă, dar nu obligatorie. Cogn. Afectează. Comportă-te. Neuroști. 8, 132–142. doi: 10.3758 / cabn.8.2.132
PubMed Abstract | CrossRef Full Text/Google Scholar
Scott, J. P. R., Mcnaughton, L. R., și Polman, R. C. J. (2006). Efectele privării de somn și a exercițiilor fizice asupra performanței cognitive, motorii și a dispoziției. Physiol. Comportă-te. 87:408. doi: 10.1016 / j. physbeh.2005.11.009
PubMed Abstract | CrossRef Full Text/Google Scholar
Smallwood, J., Brown, K., Baird, B. și Schooler, J. W. (2011). Cooperarea dintre rețeaua de mod implicit și rețeaua frontal-parietală în producerea unui tren intern de gândire. Brain Res. 1428, 60-70. doi: 10.1016 / j. brainres.2011.03.072
PubMed Abstract | CrossRef Full Text/Google Scholar
Smith, me, Mcevoy, LK, și Gevins, A. (2002). Impactul pierderii moderate a somnului asupra semnalelor neurofiziologice în timpul performanței sarcinii de memorie de lucru. Somnul 25, 784-794. doi: 10.1093/somn / 25.7.56
CrossRef Full Text/Google Scholar
Thomas, M. (2003). Baza neuronală a vigilenței și a deficiențelor de performanță cognitivă în timpul somnolenței ii. efectele 48 și 72 de ore de privare de somn asupra trezirii activității cerebrale regionale umane. Thalam. Relat. Syst. 2, 199–229. doi: 10.1016 / S1472-9288 (03) 00020-7
CrossRef Full Text | Google Scholar
Van Dongen, H. (2004). Costul cumulat al vegherii suplimentare: efectele doseresponse asupra funcțiilor neurocomportamentale și fiziologia somnului din privarea cronică de somn și privarea totală de somn. Somnul 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef Full Text | Google Scholar
Verweij, I. M., Romeijn, N., Smit, DJ și Giovanni, P. (2014). Privarea de somn duce la pierderea conectivității funcționale în regiunile frontale ale creierului. BMC Neurosci. 15:88. doi: 10.1186 / 1471-2202-15-88
PubMed Abstract | CrossRef Full Text/Google Scholar
Wen, X., Liu, Y., Yao, L. și Ding, M. (2013). Reglarea de sus în jos a activității modului implicit în atenția vizuală spațială. J. Neurosci. 33, 6444–6453. doi: 10.1523/JNEUROSCI.4939-12.2013
CrossRef Full Text | Google Scholar
Wiggins, E., Malayka, M. și Kendra, G. (2018). Privarea de somn de 24 de ore afectează modularea atențională timpurie a procesării neuronale: un studiu potențial cerebral legat de evenimente. Neuroști. Let. 677, 32–36. doi: 10.1016 / j. neulet.2018.04.022
PubMed Abstract | CrossRef Full Text/Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y. și Zhou, Q. (2019). Scăderea informațiilor înlocuirea memoriei de lucru după privarea de somn: dovezi dintr-un studiu potențial legat de evenimente. În față. Neuroști. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef Full Text/Google Scholar
Zhang, Q., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z. și colab. (2014). Un studiu ERP vizual al inhibării impulsurilor în urma unui pui de somn indus de zaleplon după privarea de somn. PLoS Unul 9: e95653. doi: 10.1371/jurnal.pone.0095653
PubMed rezumat / CrossRef textul integral / Google Scholar
Leave a Reply