Frontiers in Neuroscience
Introduction
With the progress of society and changes in work rhythm, an increasing number of people are suffering from sleep deprivation. A privação do sono não só prejudica a saúde física e mental do indivíduo, como também afeta seriamente o desempenho do trabalho, causando erros de trabalho e até mesmo acidentes. Portanto, compreender o mecanismo de privação do sono que afeta a função cognitiva é de grande significado para prevenir efetivamente os efeitos da privação do sono.estudos anteriores revelaram que a privação do sono pode causar uma série de alterações no humor, na capacidade cognitiva, no desempenho do trabalho e na função imunitária (Choo et al., 2005). A falta de sono perturba a circulação do corpo e afeta as habilidades cognitivas e emocionais dos indivíduos (Raymond, 1988). Vários estudos revelaram que a privação do sono prejudica a inibição da resposta (Harrison e Horne, 1998; Muzur et al., 2002; Jennings et al., 2003). Por exemplo, após 36 H de privação do sono, a capacidade do indivíduo de suprimir estímulos negativos diminuiu (Chuah et al., 2006). Estudos de Neuroimaging têm sugerido que a privação do sono reduz o baixo nível de capacidade de processamento visual de um indivíduo (Anderson e Platten, 2011; Ning et al., 2014). Além disso, a privação do sono prejudica o hipocampo e pode afetar a memória, destruindo a plasticidade sináptica (Cote et al., 2014). Thomas (2003) has indicated that lack of sleep reduced cerebral blood flow and metabolic rate in the thalamus, prefrontal cortex, and parietal cortex (Géraldine et al., 2005). Jarraya e colegas descobriram que a privação parcial do sono afetou significativamente as funções neuropsicológicas, tais como memória instantânea verbal, atenção e alerta (Thomas, 2003). Além disso, alguns estudos revelaram que os efeitos cumulativos da privação parcial do sono podem prejudicar gravemente a função cognitiva e o comportamento (Van Dongen, 2004; Scott et al., 2006; Jarraya et al., 2013).
memória de trabalho é um sistema usado para armazenar e processar informações e que é uma função cognitiva com capacidade limitada (Bartel et al., 2004). Além disso, a informação armazenada no sistema de memória de trabalho pode ser alterada de memória de curto prazo para memória de longo prazo através de recontagem e outros métodos de memória. A memória de trabalho é a transição entre sistemas de memória de curto e longo prazo, que é muito importante no processamento de mensagens humanas (Miyake e Shah, 1999). Ele fornece um espaço de armazenamento temporário e os recursos necessários para processar informações, tais como compreensão de voz, raciocínio e aprendizagem. A privação do sono demonstrou afectar primeiro a memória de trabalho.estudos anteriores utilizaram o paradigma da memória de N-back em participantes que sofreram privação de sono e descobriram que a falta de sono induz uma diminuição da atividade metabólica na rede regional do cérebro, que é principalmente realizada processamento de informação e inibição de reação (Baddeley, 2000; Zhang et al., 2019). Memória de trabalho deficiente após a privação do sono está relacionada com a ativação da rede padrão em tarefas (Chee e Chuah, 2008), que pode estar relacionada com o importante papel do tálamo na vigilância cortical. Por exemplo, a privação do sono aumentou a conexão entre o hipocampo, tálamo e a rede padrão, que muitas vezes era acompanhada por maior sonolência subjetiva e pior desempenho da memória de Trabalho (Lei et al., 2015; Li et al., 2016). Estudos sobre privação de sono identi?o aumento da latência e da amplitude reduzida do componente P3 foram associados a sobriedade prolongada (Morris et al., 1992; Jones and Harrison, 2001; Panjwani et al., 2010). A diminuição da onda P3 pode reflectir uma diminuição da atenção dos participantes e uma redução do discernimento dos estímulos-alvo (Koslowsky e Babkoff, 1992).no entanto, poucos estudos forneceram provas electrofisiológicas para a diminuição da memória de trabalho após a privação do sono. A tarefa N-back é considerada um método comum para avaliar a memória de trabalho (Owen et al., 2005; Jaeggi et al., 2010). Zhang et al. projetou uma tarefa de memória de trabalho de duas costas para explorar a mensagem diminuída alternativa de memória de trabalho durante a privação de sono, mas poucos estudos têm usado diferentes tipos de tarefas de memória de trabalho em uma única experiência. No presente estudo, projetamos diferentes tipos de tarefas de memória de trabalho (memória de trabalho de pronúncia, memória de trabalho espacial e memória de trabalho de objeto) para explorar o comprometimento da função cognitiva por TSD e os dados EEG do participante gravado em 2 pontos de tempo (linha de base e 36 h-TSD). Todas as tarefas adotaram um paradigma 2-back. Este estudo avaliou as mudanças na onda N2-P3 relacionadas à memória de trabalho durante a TSD e analisou as características temporais dos efeitos da privação do sono na memória de trabalho. As nossas descobertas fornecem provas experimentais dos efeitos da privação do sono na função cognitiva.
materiais e métodos
participantes
dezasseis jovens, saudáveis, destros estudantes do sexo masculino participaram neste estudo. Recrutamos participantes através de publicidade no campus. Todos os participantes tinham bons hábitos de sono (PSQI<5). Todos os participantes tinham entre 21 e 28 anos, com uma idade média de 23 anos, e nenhum dos participantes tinha qualquer doença mental ou física. Todos os participantes tinham visão normal ou visão corrigida acima de 1.0 e pontuações de inteligência > 110 no teste do Corvo. Antes da experiência, o experimentador explicou o procedimento e aponta para atenção aos participantes para garantir que eles estavam familiarizados com o método e procedimento. Nas 2 semanas anteriores à experiência, os participantes dormiam regularmente durante 7-9 h por dia, sem fumar, beber café, beber álcool, ou consumir qualquer medicação durante 2 dias antes da experiência. Antes da experiência, todos os participantes deram o seu consentimento por escrito. O esquema experimental foi aprovado pelo Comitê de Ética da quarta Universidade de Medicina militar e da Universidade Beihang.
design Experimental
três tipos de tarefas de memória de trabalho foram apresentados a todos os participantes. Eram tarefas de memória de trabalho de duas-costas (ver Figura 1), de memória de trabalho espacial de duas-costas (ver Figura 2) e de memória de trabalho de objetos de duas-costas (ver Figura 3). Os materiais estimulantes das tarefas foram 15 letras inglesas insensíveis a maiúsculas que excluindo as com letras similares, tais como L/l, M/m; pequenos quadrados pretos; e 12 figuras geométricas, respectivamente. Todos os materiais foram mostrados em preto sobre um fundo branco, com um ângulo visual aproximado de 1,5° × 1,5° (largura: 2,0 cm, Altura: 2,0 cm) subtenente. 122 trilhas foram compostas em cada tarefa e, em cada trilha, o estímulo alvo foi apresentado para 400 ms duas trilhas após a apresentação do estímulo objetivo, com o início do estímulo de 1.600 ms tempo de assíncronia (SOA) que foi marcado por um branco “+.”Os participantes foram convidados a clicar no botão esquerdo do mouse quando o alvo e o estímulo objetivo eram os mesmos (“correspondência”), enquanto clicar no botão direito do mouse quando eles não estavam (“inadequação”). A condição correspondente ou não foi apresentada numa ordem pseudorandom com uma razão 1:1.
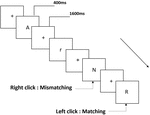
Figura 1. Diagrama esquemático da tarefa de memória de trabalho de pronúncia.
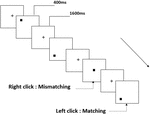
Figura 2. Diagrama esquemático da tarefa de memória de trabalho espacial.
Figura 3. Diagrama esquemático da tarefa de memória de trabalho do objecto.antes da experiência, os participantes foram instruídos sobre a tarefa experimental. Eles foram informados para praticar os três tipos de tarefas de memória de trabalho até que uma taxa de precisão de 90% foi alcançada. Os participantes visitaram o laboratório no dia anterior à experiência e dormiram no laboratório naquela noite. Os dois participantes realizaram as experiências ao mesmo tempo. Três tipos de tarefas de memória de trabalho foram realizadas às 7:30 da manhã às 8:30 da manhã seguinte com Gravação Simultânea de eletroencefalograma (EEG) (linha de base). A segunda gravação do EEG (36 h-TSD) foi realizada após um período de 36-h durante o qual os participantes não foram autorizados a dormir. Durante todo o tempo da experiência, a inibição central e drogas estimulantes foram proibidas. Os participantes foram acompanhados, observados e lembrados por enfermeiros, a fim de mantê-los acordados durante toda a sessão de TSD.
gravações EEG
um EEG de couro cabeludo contínuo foi gravado usando eletrodos colocados em 64 locais usando o sistema 10-20 com um amplificador SynAmps2. Os mastóides bilaterais (A1 e A2) foram usados como referência, e a testa foi usada como um solo. Os EEGs foram registrados a 1.000 Hz, e a impedância de todos os canais foi mantida abaixo de 5 kΩ. Quatro eletrodos adicionais foram colocados acima e abaixo dos olhos direito e esquerdo para registrar um eletrooculograma bipolar vertical e horizontal.
Análise de Dados de Experimentos Comportamentais
Devido a erros técnicos, dois casos foram eliminados, enquanto outros 14 casos foram incluídos na seguinte análise estatística. Os dados comportamentais incluíam o tempo médio de reação, a taxa correta e o número correto por unidade de tempo. Dados comportamentais na linha de base e 36 estados h-TSD foram registrados para análise. As análises foram realizadas pela IBM SPSS (V22.2), em que foi lançada a análise pós-hoc das medidas repetidas do método de variância (ANOVA) com o Geisser com efeito de estufa. Os resultados estatísticos foram apresentados como o desvio médio e padrão (DP).
análise de dados EEG
Scan 4.3 programa foi usado para analisar os dados EEG, onde os artefatos EEG do movimento ocular foram corrigidos pelo método de redução de artefatos oculares. Epochs variando de -100 a 800 ms dos dados contínuos de EEG foram extraídos e filtrados por um filtro de bandpass de 0,5 a 30 Hz com o declive de frequência de 24 dB/oct. Os ensaios em que a tensão excedeu ± 100 µV foram rejeitados e o valor inicial foi corrigido para uma amplitude média de 100 ms. os componentes do EEG foram calculados em média e calculados apenas com as respostas corrigidas. Os componentes ERP P2 (100-250 ms), N2 (150-350 ms) e P3 (250-450 ms) dos ensaios de estímulo foram identificados e quantificados. As amplitudes de Pico grand-average e latências dos componentes N2 e P3 foram calculadas separadamente em F3, Fz, F4, C3, CZ, C4, P3, Pz, e P4, e o componente P2 foi calculado em F3, Fz, F4, C3, Cz, E C4 (Casement et al., 2006; Verweij et al., 2014).medidas repetidas foi utilizada ANOVA para todos os resultados do ERP. Os efeitos principais e as interações entre os estados de sono (linha de base e 36 h-TSD), tarefas (pronúncia memória de trabalho, espacial, memória de trabalho, e o objeto de memória de trabalho), regiões (frontal, central, parietal; o componente P2 foi analisada somente no frontal e regiões centrais), e sites de esquerda, meio e direita) foram analisados estatisticamente empregando medidas repetidas ANOVA, que incluiu o Greenhouse-Geisser correções para não-esfericidade e de Bonferroni testes post-hoc.
resultados
desempenho Comportamental
os resultados dos experimentos comportamentais são mostrados na Tabela 1. O tempo médio de reacção foi mais longo no estado de 36 H-TSD do que no início, com uma tendência para aumentar, mas sem diferenças significativas. A ANOVA revelou que a taxa correcta da tarefa era significativamente diferente entre a linha de base e 36 H-TSD . O número correto por unidade de tempo mostrou um efeito principal significativo do tempo durante 36 H-TSD .

Tabela 1. Dados de desempenho (média ± DP) relativos à tarefa 2-back na linha de base e após 36 H-TSD.
Amplitude
em comparação com a linha de base, observou-se uma diminuição significativa na amplitude de P3 , e observou-se um aumento significativo na amplitude de P2 após a TSD. Embora a amplitude N2 tenha diminuído após 36 h de TSD, a diferença não atingiu significância estatística (Tabela 2).

Tabela 2. Amplitude de Pico Grand-média dos componentes P2, N2 e P3 na condição de resposta correta em vários locais de eletrodos na linha de base e após 36 h-TSD.
os principais efeitos significativos das regiões e locais na amplitude de P2 foram encontrados sob a condição TSD. Durante a TSD, a amplitude máxima de P2 apareceu na região frontal (Figura 4). Além disso, as diferenças nas amplitudes P2 em diferentes regiões (frontal vs. central) foram mais significativas nos eléctrodos bilaterais (esquerda: P = 0, 001; direita: P = 0, 1000) do que nos eléctrodos médios (Figura 4). Um efeito principal significativo da região e Local na N2 revelou que a amplitude da N2 era mais negativa no frontal do que na região central (P = 0, 008, Figura 4) e era menor à direita do que no lado esquerdo (P = 0,011, figuras 5A2, B2). Observou-se um efeito principal do local na amplitude P3 . A amplitude de P3 foi mais positiva no meio do que no lado esquerdo (P = 0, 009, Figura 4). Observou-se um efeito de interacção significativo entre o tempo e a região para a amplitude de P3 . Durante a TSD, a redução da amplitude P3 foi mais significativa nas regiões frontal e central do que na região parietal (P = 0, 005; P = 0, 003) (figura 5C3). Nenhum outro efeito principal ou efeitos de interacção atingiu significado estatístico.
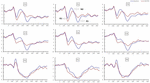
Figura 4. Amplitude do ERP na linha de base e 36 H-TSD para a condição de resposta correta para a tarefa de memória de trabalho. Os canais são ordenados da esquerda para a direita e de cima para baixo da seguinte forma: F3, Fz, F4, C3, Cz, C4, P3, Pz e P4. Em comparação com a linha de base, as latências dos componentes N2-P3 foram prolongadas e as amplitudes da N2-P3 diminuíram após 36 H-TSD.
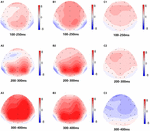
Figura 5. Mapa topográfico da resposta correta na tarefa de memória de trabalho em diferentes condições de sono (A1–C3). (A1) P2, 100-250 ms, no início. A2) N2, 200-300 ms, na linha de base. (A3) P3, 300-400 ms, na linha de base. (B1) P2, 100-250 ms, a 36 h-TSD. (B2) N2, 200-300 ms, para 36 H-TSD. (B3) P3, 300-400 ms, para 36 H-TSD. (C1) P2, 100-250 ms, 36 H-TSD com a linha de base subtraída. (C2) N2, 200-300 ms, 36 H-TSD com subtracção da linha de base. (C3) P3, 300-400 ms, 36 h-TSD com subtracção da linha de base.
latência
as latências de N2 e P2 foram significativamente prolongadas após a TSD. Embora a latência P3 tenha sido prolongada após 36 h de TSD, a diferença não atingiu significância estatística (Tabela 3).

Tabela 3. Latência de Pico Grand-média dos componentes P2, N2 e P3 na condição de resposta correta em vários locais de eletrodos na linha de base e após 36 H-TSD.
O significativo efeito principal da região em N2 e P3 revelou que a latência de N2-P3 componentes foi menor na região parietal do que na região frontal (P = 0,002; P = 0,000) e região central (P = 0,000; P = 0,000) (Figura 4). A latência da onda P3 foi significativamente mais longa no lado esquerdo do que no lado direito (Figura 4).nenhum outro efeito principal ou efeitos de interacção atingiu significado estatístico.as amplitudes e latências N2, P2 e P3 que foram elicitadas nos nove locais de eletrodos são apresentadas na Figura 4. O mapa topográfico da resposta correta na tarefa de memória de trabalho em diferentes condições de sono (linha de base, 36 h-TSD e a diferença entre as duas condições) é apresentado na Figura 5.
discussão
neste estudo, nós relatamos as influências da privação de sono de 36 h na memória de trabalho, combinando dados comportamentais em dois estados de sono (baseline e 36 H-TSD) com gravações contemporâneas de EEG. A análise dos resultados indicou que as mudanças nos dados comportamentais de acordo com a memória de trabalho deficiente após 36 H de TSD: um aumento no tempo médio de reação das tarefas cognitivas e uma diminuição na precisão.a privação do sono prejudicou o controlo individual dos recursos de atenção. Embora as pessoas tenham tentado manter a vigília e o desempenho do trabalho, incluindo o tempo de reação e a taxa correta, durante a privação do sono, a capacidade de processamento da informação de sua memória de trabalho ainda foi afetada devido à diminuição da velocidade de processamento da informação (Casement et al., 2006; Wiggins et al., 2018). Neste estudo, as ondas N2 e P3 relacionadas com a memória de trabalho foram medidas para mostrar um aumento na latência e uma diminuição na amplitude após a privação do sono em comparação com as leituras de base. Estudos têm demonstrado que a privação do sono leva a um declínio contínuo na atenção, e o fenômeno da diminuição da amplitude de P3 indica que o controle de cognição de cima para baixo dos indivíduos colapsa gradualmente. A privação do sono tem um efeito mais adverso sobre as funções cognitivas, especialmente aquelas que dependem de mental ou cognições (Kusztor et al., 2019).
O componente P3 reflete a implantação de recursos de atenção, e a latência de P3 é amplamente vista como a janela de tempo para a categorização de estímulo e avaliação. A diminuição da amplitude da onda P3 também confirmou que a tomada de decisão na resposta correspondente após a TSD tinha sido danificada em certa medida (Gosselin et al., 2005). Estudos têm sugerido que a privação do sono pode afetar a fase de processamento da Informação da memória de trabalho. Neste estudo, os indicadores de desempenho também apoiaram a conclusão de que o tempo de resposta ao estímulo alvo foi aumentado e que a latência da onda P3 foi prolongada (Cote et al., 2008). Especulou-se que o efeito da privação do sono nos componentes P3 também poderia ocorrer devido à falha em responder à alteração de informação, o que é consistente com as conclusões anteriores de que os componentes P3 estão relacionados com a atualização do conteúdo de memória de trabalho (Donchin e Fabiani, 1991).estudos anteriores consideraram o componente N2 como um índice electrofisiológico refletindo a capacidade do indivíduo de suprimir a resposta (Kreusch et al., 2014). Após a privação do sono, a latência prolongada do componente Nogo-N2 indica que a capacidade do indivíduo para suprimir a resposta é prejudicada (Jin et al., 2015). A diminuição da amplitude e latência prolongada do componente N2 relacionado com a pronúncia da memória de trabalho após a privação do sono revela que a privação do sono prejudica o processamento de informação da memória de trabalho da pronúncia (Zhang et al., 2019). O componente N2 é geralmente pensado para refletir a atenção seletiva do cérebro e o processamento de estímulos emocionais ou sinais (Schacht et al., 2008) e é um componente endógeno relacionado ao estado mental, atenção e grau de atenção de um indivíduo. Neste estudo, descobrimos que a latência do componente N2 foi significativamente prolongada, mas a amplitude mostrou apenas uma tendência descendente. De acordo com estudos anteriores, latência N2 prolongada reflectiu um aumento no tempo de resposta após restrição do sono (Zhang et al., 2014). No entanto, a descoberta de que a amplitude de N2 não foi significativamente alterada pode ter sido devido a respostas compensatórias cerebrais (Drummond e Brown, 2001). No caso de recursos cognitivos limitados, havia um mecanismo de compensação para restaurar a função cognitiva comprometida (Jin et al., 2015).
de acordo com a topografia do couro cabeludo, as alterações nos componentes N2-P3 relacionadas com a privação do sono são mais óbvias na área frontal. O controle Frontoparietal (FPC) desempenha um papel importante no controle cognitivo. Estudos têm mostrado que a FPC pode contornar o controle cognitivo top-down, permitindo que os indivíduos se concentrem em informações relacionadas com o alvo, enquanto suprimem informações que não estão relacionadas com o alvo (Smallwood et al., 2011; Wen et al., 2013). A CQP é importante para a retenção de informações e o processamento de informações na memória de trabalho, e o grau de ativação da CQP após a privação do sono foi reduzido em comparação com um grupo de sono normal (Ma et al., 2014). Embora os resultados do EEG não tenham refletido em detalhe as mudanças em regiões cerebrais específicas, intuitivamente refletiu o efeito da TSD na retenção e processamento da informação de memória de trabalho.
Embora a exata processo cognitivo que o componente P2 subjacente ainda é amplamente debatida, como uma definição ampla, o componente P2 reflete o processo de atenção e processamento visual e é, geralmente, consideradas relacionadas com a atenção seletiva e a memória de trabalho, refletindo o início do julgamento do processo perceptual (Saito et al., 2001). Neste estudo, encontramos um aumento significativo na amplitude da onda P2 após a privação do sono. Estudos têm relatado que as ondas P2 que podem ser uma parte do sistema de correspondência cognitiva inicial para processamento de mensagens e podem comparar entradas sensoriais para memória armazenada (Freunberger et al., 2007) são sensíveis a alterações na atenção da missão e demandas de memória de trabalho (Smith et al., 2002). A compensação funcional é uma das funções únicas do cérebro humano e um fator importante para a manutenção da função cognitiva. A activação excessiva do córtex pré-frontal dorsolateral (DLPFC) após a privação do sono indica que, à medida que os recursos cerebrais diminuem, o DLPFC parece ter uma função compensatória (Drummond et al., 2004; Choo et al., 2005). Portanto, especula-se que o aumento significativo da amplitude de P2 observado neste estudo pode ser devido à compensação funcional em que os indivíduos parecem manter a função cognitiva normal após a privação do sono. Embora um grande número de estudos tenham usado a tecnologia ERP para explorar o efeito da privação do sono nas funções cognitivas, componentes precoces como N1 e P2 não foram sistematicamente estudados, e os resultados são inconsistentes (Evans e Federmeier, 2007; Wiggins et al., 2018; Zhang et al., 2019). Existem poucas pesquisas sobre a mudança do componente P2 durante a privação do sono (Mograss et al., 2009). Por conseguinte, os efeitos da privação do sono nos componentes precoces do ERP, como o P2, ainda precisam de ser estudados e explorados.
Neste experimento, nós usamos o modelo 2-back para projetar a pronúncia, espacial e tarefas de memória de trabalho de objetos e examinamos o comprometimento da memória de trabalho após 36 H de TSD. Em comparação com estudos anteriores, que incidiu apenas sobre os efeitos da privação de sono em um tipo específico de informações, tais como a pronúncia de memória de trabalho, ou específicos da função cognitiva, como resposta de inibição, temos considerado o conteúdo da memória de trabalho de modelo e exaustivamente analisados os efeitos da privação do sono na memória de trabalho.no entanto, o estudo tem algumas limitações. Em primeiro lugar, nós apenas usamos a tarefa 2-back e não comparamos o desempenho dos participantes em tarefas de memória de trabalho de diferentes dificuldades. Por conseguinte, há limitações na explicação e inferição de alterações na carga de trabalho. Em segundo lugar, apenas voluntários do sexo masculino foram utilizados no estudo, e as conclusões precisam ser avaliadas quando se estendem a voluntários do sexo feminino. Devido ao número limitado de participantes, descobrimos apenas que a amplitude da onda N2 tinha uma tendência descendente e que a latência da onda P3 tinha uma tendência prolongada. Podem obter-se resultados estáveis após o aumento do número de participantes. Em terceiro lugar, combinar nosso procedimento com o fMRI para memória de trabalho pode facilitar a interpretação dos resultados. Estudos anteriores mostraram que os biorritmos circadianos afetam o desempenho comportamental, e há diferenças individuais (Montplaisir, 1981; Lavie, 2001). Não registamos os dados do EEG ao mesmo tempo neste experimental, então a influência dos biorritmos circadianos nos resultados do teste não pode ser completamente excluída.
esta pesquisa mostrou que a capacidade de memória de trabalho foi prejudicada após a TSD e que este dano não estava associado com o conteúdo de estímulo da memória de trabalho. A falta de sono reduziu a qualidade da informação armazenada na memória, o que pode ocorrer com o processo degenerativo de atenção (Ratcliff e Van Dongen, 2018). Este estudo fornece evidências de eletrofisiologia para a compreensão do mecanismo sob a memória de trabalho deficiente após a privação do sono. É necessário prestar atenção aos efeitos adversos da diminuição da memória de trabalho causada pela privação do sono e explorar intervenções eficazes para esses danos.
Declaração de disponibilidade de dados
os conjuntos de dados gerados para este estudo estão disponíveis A pedido do autor correspondente.os estudos envolvendo participantes humanos foram revisados e aprovados pela quarta Universidade de Medicina militar da Universidade Beihang. Os pacientes / participantes deram seu consentimento informado por escrito para participar deste estudo.
contribuições dos autores
Ys desenharam as experiências. A ZP produziu os resultados e escreveu o manuscrito. CD e LZ analisaram e interpretaram os dados. JT e YS realizaram os experimentos, familiarizaram os dados e os garantes deste estudo. YB, LZ e JT contribuíram para participar na coleta de dados e revisão da literatura. Todos os autores listados leram e aprovaram o manuscrito final.esta investigação foi apoiada pela National Science Foundation of Winter Olympics Technology Plan of China sob a subvenção n. º 2019YFF0301600 e HJ20191A020135.
conflito de interesses
os autores declaram que a pesquisa foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.
Anderson, C., and Platten, C. R. (2011). A privação do sono diminui a inibição e aumenta a impulsividade para estímulos negativos. Behav. Brain Res. 217:466. doi: 10.1016 / j. bbr.2010.09.020
PubMed Abstract | CrossRef Full Text | Google Scholar
Baddeley, A. (2000). O tampão episódico: um novo componente da memória de trabalho? Trends Cogn. Ciência. 4:417. doi: 10.1016/s1364-6613(00)01538-2
CrossRef Texto Completo | Google Scholar
Bartel, P., Offermeier, W., Silva, F., e Becker, P. (2004). Atenção e memória de trabalho em anestesistas residentes após o trabalho nocturno: efeitos individuais e de grupo. Occup. Ambiente. Med. 61, 167–170. doi: 10.1136/oem.2002.006197
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Casement, M. D., Broussard, J. L., Mullington, J. M., e Prima, D. Z. (2006). The contribution of sleep to improvements in working memory scanning speed: a study of prolonged sleep restriction. Biol. Psychol. 72:212. doi: 10.1016 / j. biopsycho.2005.11.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Chee, M. W. L., and Chuah, L. Y. M. (2008). Neuroimaging funcional insights into how sleep and sleep deprivation affect memory and cognition. Moeda. Opin. Neurol. 21, 417–423. doi: 10.1097 / WCO.0b013e3283052cf7
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S., e Chee, M. W. L. (2005). Dissociação de regiões corticais modulada tanto pela carga de memória de trabalho como pela privação de sono e apenas pela privação de sono. Neuroimagem 25, 579-587. doi: 10.1016 / j. neuroimage.2004.11.029
PubMed Abstract | CrossRef Full Text | Google Scholar
Chuah, Y. M. L., Venkatraman, V., Dinges, D. F., And Chee, M. W. L. (2006). A base neural da variabilidade interindividual na eficiência inibitória após a privação do sono. J. Neurosci. 26, 7156–7162. doi: 10.1523/jneurosci.0906-06. 2006
CrossRef Full Text/Google Scholar
Cote, K. A., Milner, C. E., Osip, S. L., Baker, M. L., and Cuthbert, B. P. (2008). Excitação fisiológica e atenção durante uma semana de restrição contínua do sono. Physiol. Behav. 95, 353–364. doi: 10.1016 / j. physbeh.2008.06.016
PubMed Abstract | CrossRef Full Text | Google Scholar
Cote, K. A., Mondloch, C. J., Sergeeva, V., Taylor, M., and Semplonius, T. (2014). Impacto da privação total do sono no processamento neural comportamental de rostos emocionalmente expressivos. Expo. Brain Res. 232, 1429-1442. doi: 10.1007/s00221-013-3780-1
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Donchin, E., e Fabiani, M. (1991). O uso de potenciais relacionados com eventos cerebrais no estudo da memória: p300 é uma medida da distinção de eventos? Psychol. Facul. Publico. 7, 58–77. doi: 10.1111/sltb.12055
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Drummond, S. P.; Brown, G. G., Salamat, J. S., e Gillin, J. C. (2004). A dificuldade crescente da tarefa facilita a resposta compensatória cerebral à privação total do sono. Dorme 27, 445-451. doi: 10.1093 / sleep / 27.3.445
CrossRef Full Text/Google Scholar
Drummond, S. P. A., and Brown, G. G. (2001). Os efeitos da privação total do sono nas respostas cerebrais ao desempenho cognitivo. Neuropsicofarmacologia 25, S68-S73. doi: 10.1016/S0893-133X(01)00325-6
CrossRef Texto Completo | Google Scholar
Evans, K. M., e Federmeier, K. D. (2007). A memória que\”s direita e a memória que\” s esquerda: potenciais relacionados com eventos revelam assimetrias hemisféricas na codificação e retenção da informação verbal. Neuropsicologia 45, 1777-1790. doi: 10.1016 / j. Neuropsicologia.2006.12.014
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M., e Höller, Y. (2007). O componente Visual p2 está relacionado com o bloqueio de fase theta. Neuroci. Lett. 426, 181–186. doi: 10.1016 / j. neulet.2007.08.062
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Géraldine, R., Desgranges, B., Foret, J., e Eustache, F. (2005). As relações entre sistemas de memória e estágios de sono. J. Sleep Res. 14, 123-140. doi: 10.1111 / J. 1365-2869. 2005. 00450.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Gosselin, A., Koninck, J. D., and Campbell, K. B. (2005). Privação Total de sono e processamento de novidades: implicações para o funcionamento do lobo frontal. Clin. Neurofisiol. 116, 211–222. doi: 10.1016 / j. clinph.2004.07.033
PubMed Abstract | CrossRef Full Text | Google Scholar
Harrison, Y., and Horne, J. (1998). A perda do sono prejudica tarefas de linguagem curta e nova com um foco pré-frontal. J. Sleep Res. 7, 95-100. doi: 10.1046/J. 1365-2869. 1998. 00104.x
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J., and Meier, B. (2010). A validade simultânea da tarefa N-back como medida de memória de trabalho. Memory 18, 394-412. doi: 10.1080/09658211003702171
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., e Chamari, K. (2013). O impacto da privação parcial do sono nas variações diurnas do desempenho cognitivo em indivíduos treinados. Procedimento. Soc. Behav. Ciência. 82, 392–396. doi: 10.1016 / j. sbspro.2013.06.
CrossRef texto completo/Google Scholar
Jennings, J. R., Monk, T. H., and Molen, M. W. V. D. (2003). A privação do sono influencia alguns, mas não todos os processos de atenção de supervisão. Psychol. Ciência. 14, 473–479. doi: 10.1111 / 1467-9280. 02456
PubMed Abstract | CrossRef Full Text/Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L., and Lei, Y. (2015). Recuperação reversão do sono inibição da resposta diminuída devido à restrição do sono: evidência de um estudo de potenciais relacionados com acontecimentos visuais. PLoS 1 10:e0142361. doi: 10.1371 / journal.pone.0142361
PubMed Abstract | CrossRef Full Text | Google Scholar
Jones, K., and Harrison, Y. (2001). Função do lobo Frontal, perda de sono e sono fragmentado. Med Do Sono. AP.5:475. doi: 10.1053 / smrv.2001.0203
PubMed Abstract | CrossRef Full Text | Google Scholar
Koslowsky, M., and Babkoff, H. (1992). Meta-análise da relação entre a privação total do sono e o desempenho. Chronobiol. T. 9, 132–136. doi: 10.3109/07420529209064524
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A., e Hansenne, M. (2014). Abuso de álcool e componentes erp em tarefas go / no-go usando estímulos relacionados ao álcool: impacto da evasão de álcool. T. J. Psychophysiol. 94, 92–99. doi: 10.1016 / j. ijpsycho.2014.08.001
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Tempestade, J. F., e Huster, R. J. (2019). A privação do sono afeta subcomponentes do controle cognitivo. Sleep 42: zsz016. doi: 10.1093 / sleep / zsz016
PubMed Abstract | CrossRef Full Text/Google Scholar
Lavie, P. (2001). Sono-acordar como um ritmo biológico. Annu. Rev. Psychol. 52, 277–303. doi: 10.1146 / annurev.psicologia.52.1.277
CrossRef Texto Completo | Google Scholar
a Lei, Y., Shao, Y. Wang, L., Zhai, T., e Zou, F. (2015). O acoplamento em rede cerebral em larga escala prevê efeitos de privação total do sono na capacidade cognitiva. PLoS One 10: e0133959. doi: 10.1371 / journal.pone.0133959
PubMed Abstract | CrossRef Full Text/Google Scholar
Li, C., Huang, D., Qi, J., Chang, H., and Meng, Q. (2016). Os défices de memória a curto prazo correlacionam-se com alterações de conectividade funcional hipocampo-talâmica após restrição aguda do sono. Imagem Cerebral Do Behav. 11, 954–963. doi: 10.1007/s11682-016-9570-1
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Ma, N., Dinges, D. F., Basner, M., e Rao, H. (2014). Como a perda total aguda do sono afeta o cérebro presente: uma meta-análise de estudos de neuroimagem. Dorme 38, 233. doi: 10.5665 / sono.4404
PubMed Abstract | CrossRef Full Text | Google Scholar
Miyake, A., and Shah, P. (1999). Modelos de memória de trabalho. Mecanismos de manutenção ativa e controle executivo. Cambridge: Cambridge University Press.
Google Scholar
Mograss, M. A., Guillem, F., Brazzini-Poisson, V., and Godbout, R. (2009). Os efeitos da privação total do sono nos processos de memória de reconhecimento: um estudo do potencial relacionado com eventos. Neurobiol. Aprender. Memoria. 91, 343–352. doi: 10.1016 / j. nlm.2009.01.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Montplaisir, J. (1981). Depressão e ritmo biológico: efeitos terapêuticos da privação do sono. L’Unión Médicale du Canada 110, 272-276. doi: 10.1620 / tjem.133.481
PubMed Abstract / CrossRef Full Text | Google Scholar
Morris, A. M., So, Y., Lee, K. A., Lash, A. A., and Becker, C. E. (1992). O potencial relacionado com o evento p300. os efeitos da privação do sono. J. Occupat. Med. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F., And Hobson, J. A. (2002). O córtex pré-frontal a dormir. Trends Cogn. Ciência. 6, 475–481. doi: 10.1016/S1364-6613(02)01992-7
CrossRef Texto Completo | Google Scholar
Ning, M., Dinges, D. F., Mathias, B., e Hengyi, R. (2014). Como a perda total aguda do sono afeta o cérebro presente: uma meta-análise de estudos de neuroimagem. Dorme 38, 233-240. doi: 10.5665 / sono.4404
PubMed Abstract | CrossRef Full Text | Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, A. R., and Bullmore, E. (2005). N-back working memory paradigm: a meta-analysis of normative functional neuroimaging studies. Cantarolar. MAPP Cerebral. 25, 46–59. doi: 10.1002 / hbm.20131
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S., e Kumar, S. (2010). Os correlatos eletrofisiológicos de cognição melhoram com a nap durante a privação do sono. Euro. J. Appl. Physiol. 108, 549–556. doi: 10.1007/s00421-009-1222-3
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Ratcliff, R., e Van Dongen, H. (2018). Os efeitos da privação do sono no item e na memória de reconhecimento associativo. J. Exp. Psychol. 44, 193–208. doi: 10.1037/xlm0000452
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Raymond, C. A. (1988). Mudança de trabalho, ciclos de sono estão a caminho de se tornar outra questão de saúde pública. JAMA J. Am. Med. Assoc. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef Texto Completo | Google Scholar
Saito, H., Yamazaki, H., Matsuoka, H., Matsumoto, K., e Sato, M. (2001). Potencial relacionado com acontecimentos visuais na demência ligeira do tipo de alzheimer. Psiquiatria Clin. Neuroci. 55, 365–371. doi: 10.1046/J. 1440-1819. 2001. 00876.X
PubMed Abstract | CrossRef Full Text | Google Scholar
Schacht, A., Werheid, K., and Sommer, W. (2008). A avaliação da beleza facial é rápida, mas não obrigatória. Cogn. Afectar. Behav. Neuroci. 8, 132–142. doi: 10.3758 / cabn.8.2.132
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Scott, J. P. R., Mcnaughton, L. R., e Polman, R. C. J. (2006). Efeitos da privação do sono e do exercício sobre o desempenho cognitivo, motor e humor. Physiol. Behav. 87:408. doi: 10.1016 / j. physbeh.2005.11.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Smallwood, J., Brown, K.), Baird, B., and Schooler, J. W. (2011). Cooperação entre a rede de modo padrão e a rede frontal-parietal na produção de um trem interno de pensamento. Brain Res. 1428, 60-70. doi: 10.1016 / j. brainres.2011.03.072
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, M. E., Mcevoy, L. K., and Gevins, A. (2002). O impacto da perda moderada do sono nos sinais neurofisiológicos durante o desempenho da tarefa memória de trabalho. Dorme 25, 784-794. doi: 10.1093 / sono / 25.7.56
CrossRef Full Text | Google Scholar
Thomas, M. (2003). Base Neural de alerta e deficiências do desempenho cognitivo durante a sonolência II. efeitos de 48 e 72 horas de privação do sono na actividade cerebral regional humana em estado de vigília. Thalam. Relat. Syst. 2, 199–229. doi: 10.1016/S1472-9288(03)00020-7
CrossRef Texto Completo | Google Scholar
Van Dongen, H. (2004). O custo cumulativo da vigília adicional: efeitos de doseresponse sobre as funções neurocomportamentais e a fisiologia do sono devido à privação crónica do sono e à privação total do sono. Dorme 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef Texto Completo | Google Scholar
Verweij, I. M., Romeijn, N., Smit, D. J., e Giovanni, P. (2014). A privação do sono leva a uma perda de conectividade funcional nas regiões do cérebro frontal. Neurociência BMC. 15:88. doi: 10.1186/1471-2202-15-88
PubMed Abstract | CrossRef Full Text / Google Scholar
Wen, X., Liu, Y., Yao, L. ,and Ding, M. (2013). Regulação Top-down da actividade do modo predefinido na atenção visual espacial. J. Neurosci. 33, 6444–6453. doi: 10.1523/JNEUROSCI.4939-12. 2013
CrossRef Full Text | Google Scholar
Wiggins, E., Malayka, M., and Kendra, G. (2018). Privação de sono 24-h prejudica a modulação precoce de atenção do processamento neural: um estudo de potencial cerebral relacionado a eventos. Neuroci. Lett. 677, 32–36. doi: 10.1016 / j. neulet.2018.04.022
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y., e Zhou, P. (2019). Diminuição da informação substituição da memória de trabalho após a privação do sono: evidência de um estudo potencial relacionado com um evento. Frente. Neuroci. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef Texto Completo | Google Scholar
Zhang, P., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z. et al. (2014). Um estudo erp visual da inibição dos impulsos após uma sesta induzida pelo zaleplon após privação do sono. PLoS One 9: e95653. doi: 10.1371 / journal.pone.0095653
PubMed Abstract / CrossRef Full Text | Google Scholar
Leave a Reply