Wiązanie peptydowe
znaczna delokalizacja samotnej pary elektronów na atomie azotu nadaje grupie cząstkowy charakter wiązania podwójnego. Częściowe wiązanie podwójne powoduje powstanie grupy amidowej planarnej, występującej w izomerach cis lub trans. W stanie rozłożonym białek grupy peptydowe mogą swobodnie izomeryzować i przyjmować oba izomery; jednak w stanie złożonym tylko jeden izomer jest przyjmowany w każdej pozycji (z rzadkimi wyjątkami). Forma trans jest korzystna głównie w większości wiązań peptydowych (w przybliżeniu stosunek 1000:1 w populacjach Trans:cis). Jednak grupy peptydowe X-Pro mają zwykle stosunek około 30:1, prawdopodobnie dlatego, że symetria między c α {\displaystyle \mathrm {c^{\alpha }} }

I c δ {\displaystyle \mathrm {c^{\delta }} }

Atomy proliny sprawiają, że izomery cis i trans są prawie równe pod względem energii (patrz rysunek poniżej).
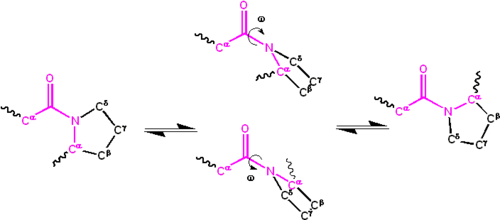
kąt dwuścienny związany z grupą peptydową (zdefiniowaną przez cztery atomy c α − c ’ − n − c α {\displaystyle C^{\alpha }-C^{\prime }-N-c^{\alpha }}

) jest oznaczony ω {\displaystyle \Omega }

; ω = 0 ∘ {\displaystyle \omega =0^{\circ }}

dla izomeru cis (konformacji synperyplanarnej) i ω = 180 ∘ {\displaystyle \omega =180^{\circ }}

dla izomeru Trans (konformacji antyperiplanarnej). Grupy amidowe mogą izomeryzować Wiązanie C ’- N pomiędzy formami cis i trans, aczkolwiek powoli ( τ ∼ {\displaystyle \tau \sim}

20 sekund w temperaturze pokojowej). Stan przejściowy ω = ± 90 ∘ {\displaystyle \ omega = \ pm 90^{\circ}}

wymaga, aby częściowe wiązanie podwójne zostało zerwane, tak że energia aktywacji wynosi około 80 kilodżula/mol (20 kcal/mol). Jednak energia aktywacji może być obniżona (i izomeryzacja katalizowana) przez zmiany, które sprzyjają postaci jednoprzęsłowej, takie jak umieszczenie grupy peptydowej w środowisku hydrofobowym lub przekazanie wiązania wodorowego do atomu azotu grupy peptydowej X-Pro. Oba te mechanizmy obniżania energii aktywacji zaobserwowano w izomerazach peptydylo-prolilowych (Ppiazach), które są naturalnie występującymi enzymami katalizującymi izomeryzację cis-trans wiązań peptydowych X-Pro.
konformacyjne składanie białek jest zwykle znacznie szybsze (Zwykle 10-100 ms) niż izomeryzacja cis-trans (10-100 s). Nieaktywny izomer niektórych grup peptydowych może znacząco zakłócić fałdowanie konformacyjne, spowalniając je lub zapobiegając nawet występowaniu, aż do osiągnięcia natywnego izomeru. Jednak nie wszystkie grupy peptydowe mają taki sam wpływ na składanie; nieaktywne izomery innych grup peptydowych mogą w ogóle nie wpływać na fałdowanie.
Leave a Reply