Frontiers in Neuroscience
wprowadzenie
wraz z postępem społeczeństwa i zmianami rytmu pracy coraz więcej osób cierpi na brak snu. Brak snu nie tylko szkodzi zdrowiu fizycznemu i psychicznemu jednostki, ale także poważnie wpływa na wydajność pracy, powodując błędy w pracy, a nawet wypadki. Dlatego zrozumienie mechanizmu deprywacji snu, który wpływa na funkcje poznawcze, ma ogromne znaczenie dla skutecznego zapobiegania skutkom deprywacji snu.
poprzednie badania wykazały, że brak snu może powodować szereg zmian nastroju, zdolności poznawczych, wydajności pracy i funkcji immunologicznych (Choo et al., 2005). Brak snu zaburza krążenie ciała i wpływa na zdolności poznawcze i emocjonalne jednostek (Raymond, 1988). Kilka badań wykazało, że brak snu upośledza hamowanie reakcji (Harrison and Horne, 1998; Muzur et al., 2002; Jennings et al., 2003). Na przykład po 36 godzinach braku snu zdolność jednostki do tłumienia negatywnych bodźców zmniejszyła się(Chuah et al., 2006). Badania neuroobrazowania sugerują, że brak snu zmniejsza niski poziom zdolności przetwarzania wizualnego osoby (Anderson and Platten, 2011; Ning et al., 2014). Ponadto brak snu upośledza hipokamp i może wpływać na pamięć poprzez niszczenie plastyczności synaptycznej(Cote et al., 2014). Thomas (2003) wskazał, że brak snu zmniejszył mózgowy przepływ krwi i tempo metabolizmu w wzgórzu, korze przedczołowej i korze ciemieniowej (Géraldine et al., 2005). Jarraya i współpracownicy odkryli, że częściowy brak snu znacząco wpływa na funkcje neuropsychologiczne, takie jak werbalna pamięć natychmiastowa, uwaga i czujność (Thomas, 2003). Ponadto niektóre badania wykazały, że skumulowane skutki częściowego braku snu mogą poważnie upośledzać funkcje poznawcze i zachowanie (Van Dongen, 2004; Scott et al., 2006; Jarraya et al., 2013).
pamięć robocza to system, który kiedyś przechowywał i przetwarzał informacje i który jest funkcją poznawczą o ograniczonej pojemności (Bartel et al., 2004). Ponadto informacje przechowywane w systemie pamięci roboczej mogą być zmieniane z pamięci krótkotrwałej na pamięć długotrwałą za pomocą retellingu i innych metod pamięci. Pamięć robocza to przejście między krótko-i długoterminowymi systemami pamięci, które jest bardzo kluczowe w przetwarzaniu wiadomości przez człowieka (Miyake and Shah, 1999). Zapewnia tymczasową przestrzeń do przechowywania i zasoby potrzebne do przetwarzania informacji, takich jak rozumienie głosu, rozumowanie i uczenie się. Wykazano, że brak snu wpłynie najpierw na pamięć roboczą.
poprzednie badania wykorzystywały paradygmat pamięci roboczej N-back u uczestników, którzy poddali się deprywacji snu i odkryli, że brak snu powoduje zmniejszenie aktywności metabolicznej w sieci regionalnej mózgu, co odbywa się głównie w przetwarzaniu informacji i hamowaniu reakcji (Baddeley, 2000; Zhang et al., 2019). Upośledzenie pamięci roboczej po deprywacji snu jest związane z aktywacją sieci domyślnej w zadaniach (Chee and Chuah, 2008), co może być związane z ważną rolą wzgórza w czujności korowej. Na przykład brak snu zwiększył połączenie między hipokampem, wzgórzem i siecią domyślną, której często towarzyszyła wyższa subiektywna senność i gorsza wydajność pamięci roboczej (Lei et al., 2015; Li et al., 2016). Badania nad brakiem snu?ze zwiększone opóźnienie i zmniejszona Amplituda składnika P3 były związane z przedłużoną trzeźwością (Morris et al., 1992; Jones and Harrison, 2001; Panjwani et al., 2010). Spadek fali P3 może odzwierciedlać spadek uwagi uczestników i zmniejszenie rozpoznawania bodźców docelowych (Koslowsky and Babkoff, 1992).
jednak niewiele badań dostarczyło dowodów elektrofizjologicznych na upośledzenie pamięci roboczej po braku snu. Zadanie N-back jest uważana za wspólną metodę oceny pamięci roboczej (Owen et al., 2005; Jaeggi et al., 2010). Zhang et al. zaprojektowano zadanie pamięci roboczej wymowy z dwoma plecami, aby zbadać zmniejszoną alternatywę wiadomości pamięci roboczej podczas braku snu, ale niewiele badań wykorzystało różne rodzaje zadań pamięci roboczej w jednym eksperymencie. W niniejszym opracowaniu zaprojektowaliśmy różne typy zadań pamięci roboczej (pamięć robocza wymowy, pamięć robocza przestrzenna i pamięć robocza obiektu) w celu zbadania upośledzenia funkcji poznawczych przez TSD i zarejestrowania danych EEG uczestnika w 2 punktach czasowych (linia bazowa i 36 H-TSD). Wszystkie zadania przyjęły paradygmat 2-tylny. W badaniu oceniono zmiany w fali N2-P3 związane z pamięcią roboczą podczas TSD i przeanalizowano czasową charakterystykę wpływu braku snu na pamięć roboczą. Nasze odkrycia dostarczają eksperymentalnych dowodów na wpływ braku snu na funkcje poznawcze.
materiały i metody
uczestnicy
w badaniu wzięło udział szesnastu młodych, zdrowych, praworęcznych mężczyzn. Rekrutowaliśmy uczestników poprzez reklamę na kampusie. Wszyscy uczestnicy mieli dobre nawyki senne (PSQI<5). Wszyscy uczestnicy byli w wieku od 21 do 28 lat ze średnią wieku 23 lat, a żaden z uczestników nie miał żadnej choroby psychicznej lub fizycznej. Wszyscy uczestnicy mieli normalne lub poprawione widzenie powyżej 1,0 i wynik inteligencji >110 W teście Raven. Przed eksperymentem eksperymentator wyjaśnił procedurę i zwraca uwagę uczestnikom, aby upewnić się, że byli zaznajomieni z metodą i procedurą. W ciągu 2 tygodni przed eksperymentem uczestnicy spali regularnie przez 7-9 godzin dziennie, bez palenia, picia kawy, picia alkoholu lub spożywania jakichkolwiek leków przez 2 dni przed eksperymentem. Przed eksperymentem wszyscy uczestnicy udzielili pisemnej świadomej zgody. Program eksperymentalny został zatwierdzony przez Komisję Etyki czwartego Wojskowego Uniwersytetu Medycznego i Uniwersytetu Beihang.
projekt eksperymentalny
wszystkim uczestnikom przedstawiono trzy rodzaje zadań pamięci roboczej. Były to zadania z pamięcią roboczą z dwoma tylnymi wymowami (patrz rysunek 1), dwa tylne zadania z pamięcią roboczą przestrzenną (patrz rysunek 2) i dwa tylne zadania z pamięcią roboczą obiektu (patrz rysunek 3). Materiały do zadań stanowiły 15 liter angielskich bez rozróżniania wielkości liter, z wyjątkiem tych o podobnych literach, takich jak L/l, M/M, małe czarne kwadraty oraz 12 figur geometrycznych. Wszystkie materiały zostały przedstawione w kolorze czarnym na białym tle, o przybliżonym kącie widzenia 1,5° × 1,5° (szerokość: 2,0 cm, Wysokość: 2,0 cm). W każdym zadaniu uwzględniono 122 szlaki, a na każdym szlaku zaprezentowano bodziec docelowy dla 400 ms dwa szlaki po prezentacji bodźca obiektywnego, z czasem asynchronicznym wystąpienia bodźca 1,600 ms (SOA) oznaczonym białym „+.”Uczestnicy zostali poproszeni o kliknięcie lewym przyciskiem myszy, gdy cel i cel były takie same („dopasowanie”), natomiast kliknięcie prawym przyciskiem myszy, gdy nie były („niedopasowanie”). Warunek pasujący lub nie został przedstawiony w kolejności pseudorandomowej w stosunku 1: 1.
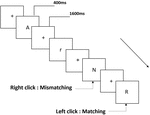
Rysunek 1. Schemat zadania pamięci roboczej wymowy.
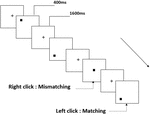
Rysunek 2. Schemat zadania przestrzennej pamięci roboczej.
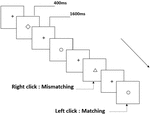
Rysunek 3. Schemat zadania pamięci roboczej obiektu.
procedury eksperymentalne
przed rozpoczęciem eksperymentu uczestnicy zostali poinstruowani o zadaniu eksperymentalnym. Poinformowano ich, aby ćwiczyli trzy rodzaje zadań pamięci roboczej, aż do osiągnięcia wskaźnika dokładności 90%. Uczestnicy odwiedzili laboratorium dzień przed eksperymentem i spali w laboratorium tej nocy. Obaj partnerzy przeprowadzili eksperymenty w tym samym czasie. W godzinach od 7:30 do 8:30 następnego dnia rano wykonywano trzy rodzaje zadań z pamięcią roboczą z równoczesnym zapisem elektroencefalogramu (EEG) (linia bazowa). Drugie nagranie EEG (36 h-TSD) zostało przeprowadzone po 36-godzinnym okresie, podczas którego uczestnicy nie mogli spać. Przez cały czas eksperymentu zabronione było Centralne hamowanie i leki pobudzające. Uczestnikom towarzyszył, obserwował i przypominał personel pielęgniarski, aby nie zasnąć przez całą sesję TSD.
nagrania EEG
ciągłe EEG zostało zarejestrowane za pomocą nakładek elektrodowych umieszczonych w 64 miejscach przy użyciu systemu 10-20 ze wzmacniaczem SynAmps2. Obustronne mastoidy (A1 i A2) były używane jako odniesienia, a czoło było używane jako podłoże. EEG rejestrowano przy 1000 Hz, a impedancja wszystkich kanałów utrzymywała się poniżej 5 kΩ. Cztery dodatkowe elektrody zostały umieszczone nad i pod prawym I lewym okiem, aby zarejestrować bipolarny pionowy i poziomy elektrookulogram.
Analiza danych eksperymentów behawioralnych
z powodu błędów technicznych dwa przypadki zostały usunięte, a pozostałe 14 przypadków włączono do poniższej analizy statystycznej. Dane behawioralne obejmowały średni czas reakcji, prawidłową szybkość i prawidłową liczbę na jednostkę czasu. Do analizy zarejestrowano dane behawioralne w Stanach wyjściowych i 36 h-TSD. Analizy zostały przeprowadzone przez IBM SPSS (V22.2), gdzie rozpoczęto analizę powtarzających się pomiarów wariancji (ANOVA) metodą Greenhouse-Geissera z analizą post-hoc. Wyniki statystyczne przedstawiono jako średnią i odchylenie standardowe (SD).
Analiza danych EEG
program Scan 4.3 został użyty do analizy danych EEG, gdzie artefakty EEG ruchu oka zostały skorygowane metodą redukcji artefaktów ocznych. Okresy od -100 do 800 ms ciągłych danych EEG były wyodrębniane i filtrowane za pomocą filtra pasmowego od 0,5 do 30 Hz o nachyleniu częstotliwości 24 dB/oct. Próby, w których napięcie przekraczało ± 100 µV, odrzucono, a wartość wyjściową skorygowano do średniej amplitudy 100 ms. komponenty EEG uśredniono i obliczono z uwzględnieniem tylko skorygowanych odpowiedzi. Składniki ERP P2 (100-250 ms), N2 (150-350 ms) i P3 (250-450 ms) w badaniach stymulacyjnych zostały zidentyfikowane i ocenione ilościowo. Grand-average peak amplitudes and latencies of the N2 and P3 components was calculated separate at F3, Fz, F4, C3, Cz, C4, P3, Pz, and P4, and the P2 component was calculated at F3, Fz, F4, C3, Cz, and C4 (Casement et al., 2006; Verweij et al., 2014).
dla wszystkich wyników ERP zastosowano powtarzalne pomiary ANOVA. Główne efekty i interakcje między Stanami snu (linia bazowa i 36 H-TSD), zadania (pamięć robocza wymowy, pamięć robocza przestrzenna i pamięć robocza obiektu), regiony (przedni, Centralny i ciemieniowy; Składnik P2 analizowano tylko na obszarach przednich i centralnych), a miejsca (lewy, środkowy i prawy) były analizowane statystycznie przy użyciu powtarzanych pomiarów ANOVA, które obejmowały poprawki Greenhouse-Geissera dla testów post-hoc non-sferyczność i Bonferroni.
wyniki
wydajność behawioralna
wyniki eksperymentów behawioralnych przedstawiono w tabeli 1. Średni czas reakcji był dłuższy w stanie 36 H-TSD niż na początku badania, z tendencją do zwiększania się, ale bez znaczących różnic. ANOVA ujawniła, że prawidłowa szybkość wykonania zadania była znacząco różna między wartością wyjściową a 36 H-TSD . Prawidłowa liczba na jednostkę czasu wykazała znaczący główny wpływ czasu podczas 36 H-TSD .

Tabela 1. Dane dotyczące wydajności (średnia ± SD) dla zadania 2-back na początku i po 36 h-TSD.
Amplituda
w porównaniu do wartości wyjściowych zaobserwowano istotne zmniejszenie amplitudy P3 i znaczące zwiększenie amplitudy P2 po TSD. Chociaż Amplituda N2 zmniejszyła się po 36 godzinach TSD, różnica nie osiągnęła istotności statystycznej (Tabela 2).

Tabela 2. Grand-średnia szczytowa Amplituda składników P2, N2 i P3 w Warunkach prawidłowej odpowiedzi w wielu miejscach elektrody na początku i po 36 h-TSD.
w Warunkach TSD stwierdzono znaczący wpływ regionów i miejsc na amplitudę P2. Podczas TSD maksymalna amplituda P2 pojawiła się w obszarze czołowym (ryc. 4). Ponadto różnice w amplitudach P2 w różnych regionach (przedni vs centralny) były bardziej znaczące w elektrodach obustronnych (po lewej: P = 0,001; po prawej: P = 0,000) niż w elektrodach środkowych (ryc. 4). Znaczący wpływ regionu i miejsca na N2 ujawnił, że amplituda N2 była bardziej ujemna w obszarze czołowym niż w obszarze centralnym (P = 0,008, Fig.4) i była mniejsza po prawej stronie niż po lewej stronie (P = 0,011,Fig. 5A2, B2). Zaobserwowano główny wpływ miejsca na amplitudę P3 . Amplituda P3 była bardziej dodatnia w środku niż po lewej stronie (P = 0,009, Fig. 4). Dla amplitudy P3 zaobserwowano istotny efekt interakcji pomiędzy czasem i regionem . Podczas TSD zmniejszenie amplitudy P3 było bardziej znaczące w regionie czołowym i centralnym niż w regionie ciemieniowym (P = 0,005; P = 0,003) (Fig.5C3). Żadne inne główne działania lub interakcje nie osiągnęły istotności statystycznej.
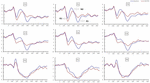
Rysunek 4. Amplituda ERP na poziomie wyjściowym i 36 h-TSD dla prawidłowego stanu odpowiedzi dla zadania pamięci roboczej. Kanały są uporządkowane od lewej do prawej i od góry do dołu w następujący sposób: F3, Fz, F4, C3, Cz, C4, P3, Pz i P4. W porównaniu do wartości wyjściowych, opóźnienia składników N2-P3 były wydłużone, a amplitudy N2-P3 były zmniejszone po 36 h-TSD.
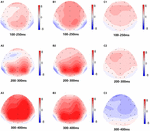
Rysunek 5. Mapa topograficzna prawidłowej odpowiedzi w zadaniu pamięci roboczej w różnych warunkach snu (A1-C3). (A1) P2, 100-250 ms, na początku. (A2) N2, 200-300 ms, na początku. (A3) P3, 300-400 ms, na początku. (B1) P2, 100-250 ms, przy 36 H-TSD. (B2) N2, 200-300 ms, dla 36 H-TSD. (B3) P3, 300-400 ms, dla 36 H-TSD. (C1) P2, 100-250 ms, 36 H-TSD z odjęciem wartości wyjściowej. (C2) N2, 200-300 ms, 36 H-TSD z odjęciem wartości wyjściowej. (C3) P3, 300-400 ms, 36 H-TSD z odjęciem wartości wyjściowej.
opóźnienie
opóźnienia N2 i P2 były znacząco wydłużone po TSD. Chociaż opóźnienie P3 było przedłużone po 36 godzinach TSD, różnica nie osiągnęła istotności statystycznej(Tabela 3).

Tabela 3. Grand-średnie opóźnienie szczytowe składników P2, N2 i P3 w Warunkach prawidłowej odpowiedzi w wielu miejscach elektrody na początku i po 36 h-TSD.
znaczący główny wpływ regionu na N2 i P3 ujawnił, że opóźnienie składników N2-P3 było krótsze w regionie ciemieniowym niż w regionie czołowym (P = 0, 002; P = 0, 000) i centralnym (P = 0, 000; P = 0, 000) (ryc. 4). Opóźnienie fali P3 było znacznie dłuższe po lewej stronie niż po prawej (ryc. 4).
żadne inne główne efekty lub efekty interakcji nie osiągnęły istotności statystycznej.
amplitudy i opóźnienia N2, P2 i P3, które zostały wywołane w dziewięciu miejscach elektrod, przedstawiono na fig.4. Mapa topograficzna prawidłowej odpowiedzi w zadaniu pamięci roboczej w różnych warunkach snu (wartość wyjściowa, 36 h-TSD i różnica między tymi dwoma warunkami) przedstawiono na fig.5.
dyskusja
w tym badaniu opisaliśmy wpływ 36-godzinnego braku snu na pamięć roboczą, łącząc dane behawioralne w dwóch stanach snu (wyjściowym i 36-H-TSD) z równoczesnymi zapisami EEG. Analiza wyników wykazała, że zmiany w danych behawioralnych zgodnie z upośledzoną pamięcią roboczą po 36 h TSD: wzrost średniego czasu reakcji zadań poznawczych i spadek dokładności.
brak snu zaburzył indywidualną kontrolę nad zasobami uwagi. Chociaż osoby starały się utrzymać czuwanie i wydajność pracy, w tym czas reakcji i prawidłową szybkość, podczas braku snu, zdolność przetwarzania informacji w ich pamięci roboczej była nadal dotknięta ze względu na spadek szybkości przetwarzania informacji (Casement et al., 2006; Wiggins et al., 2018). W tym badaniu zmierzono fale N2 i P3 związane z pamięcią roboczą w celu wykazania wzrostu opóźnienia i zmniejszenia amplitudy po braku snu w porównaniu z odczytami wyjściowymi. Badania wykazały, że brak snu prowadzi do ciągłego spadku uwagi, a zjawisko zmniejszonej amplitudy P3 wskazuje, że odgórna Kontrola funkcji poznawczych stopniowo zanika. Brak snu ma bardziej niekorzystny wpływ na funkcje poznawcze, zwłaszcza te, które zależą od funkcji umysłowych lub poznawczych (Kusztor et al., 2019).
komponent P3 odzwierciedla rozmieszczenie zasobów uwagi, a opóźnienie P3 jest powszechnie postrzegane jako okno czasowe dla kategoryzacji i oceny bodźców. Spadek amplitudy fali P3 potwierdził również, że podejmowanie decyzji w dopasowanej odpowiedzi po TSD zostało do pewnego stopnia uszkodzone(Gosselin et al., 2005). Badania sugerują, że brak snu może wpływać na etap przetwarzania informacji w pamięci roboczej. W tym badaniu wskaźniki wydajności poparły również wniosek, że czas reakcji na bodziec docelowy został zwiększony i że opóźnienie fali P3 zostało wydłużone(Cote et al., 2008). Spekulowano, że wpływ braku snu na składniki P3 może również mieć miejsce z powodu braku reakcji na zmianę informacji, co jest zgodne z wcześniejszymi wnioskami, że składniki P3 są związane z aktualizacją zawartości pamięci roboczej (Donchin and Fabiani, 1991).
poprzednie badania uznały Składnik N2 za wskaźnik elektrofizjologiczny odzwierciedlający zdolność jednostki do tłumienia odpowiedzi (Kreusch et al., 2014). Po deprywacji snu przedłużone opóźnienie składnika NoGo-N2 wskazuje, że zdolność jednostki do tłumienia odpowiedzi jest osłabiona(Jin et al., 2015). Zmniejszona amplituda i przedłużone opóźnienie komponentu N2 związane z pamięcią roboczą wymowy po deprywacji snu ujawnia, że deprywacja snu upośledza przetwarzanie informacji o pamięci roboczej wymowy (Zhang et al., 2019). Uważa się, że składnik N2 odzwierciedla selektywną uwagę mózgu i przetwarzanie bodźców lub sygnałów emocjonalnych (Schacht et al., 2008) i jest endogennym składnikiem związanym ze stanem psychicznym jednostki, uwagą i stopniem uwagi. W tym badaniu odkryliśmy, że opóźnienie składnika N2 było znacznie wydłużone, ale Amplituda wykazywała tylko tendencję spadkową. Zgodnie z wcześniejszymi badaniami wydłużone opóźnienie N2 odzwierciedlało wzrost czasu reakcji po ograniczeniu snu (Zhang et al., 2014). Jednak odkrycie, że amplituda N2 nie została znacząco zmieniona, mogło być spowodowane mózgowymi odpowiedziami kompensacyjnymi (Drummond and Brown, 2001). W przypadku ograniczonych zasobów poznawczych istniał mechanizm kompensacji w celu przywrócenia upośledzonych funkcji poznawczych (Jin et al., 2015).
zgodnie z topografią skóry głowy zmiany w składnikach N2-P3 związane z brakiem snu są bardziej widoczne w obszarze czołowym. Kontrola frontoparietalna (FPC) odgrywa ważną rolę w kontroli poznawczej. Badania wykazały, że FPC może ominąć odgórną kontrolę poznawczą, umożliwiając osobom skupienie się na informacjach związanych z celem, jednocześnie tłumiąc informacje, które nie są związane z celem (Smallwood et al., 2011; Wen et al., 2013). FPC jest ważne dla przechowywania informacji i przetwarzania informacji w pamięci roboczej, a stopień aktywacji FPC po deprywacji snu został zmniejszony w porównaniu do normalnej grupy snu (Ma et al., 2014). Chociaż wyniki EEG nie odzwierciedlały szczegółowo zmian w określonych regionach mózgu, intuicyjnie odzwierciedlały wpływ TSD na retencję i przetwarzanie informacji o pamięci roboczej.
chociaż dokładny proces poznawczy, który leży u podstaw komponentu P2, jest nadal szeroko dyskutowany, jako szeroka definicja, komponent P2 odzwierciedla proces uwagi i przetwarzania wizualnego i jest ogólnie uważany za związany z selektywną uwagą i pamięcią roboczą, odzwierciedlając wczesną ocenę procesu percepcyjnego (Saito et al., 2001). W tym badaniu stwierdziliśmy znaczny wzrost amplitudy fali P2 po braku snu. Badania donoszą, że fale P2, które mogą być częścią wczesnego poznawczego systemu dopasowywania do przetwarzania wiadomości i mogą porównywać dane sensoryczne z pamięcią przechowywaną (Freunberger et al., 2007) są wrażliwe na zmiany w misji uwagi i Wymagania pamięci roboczej (Smith et al., 2002). Kompensacja funkcjonalna jest jedną z unikalnych funkcji ludzkiego mózgu i ważnym czynnikiem utrzymania funkcji poznawczych. Nadmierna aktywacja grzbietowo-bocznej kory przedczołowej (DLPFC) po deprywacji snu wskazuje, że w miarę zmniejszania się zasobów mózgu dlpfc wydaje się mieć funkcję kompensacyjną(Drummond et al., 2004; Choo et al., 2005). Dlatego spekulujemy, że znaczny wzrost amplitudy P2 obserwowany w tym badaniu może być spowodowany kompensacją czynnościową, w której osoby wydają się utrzymywać normalne funkcje poznawcze po braku snu. Chociaż w wielu badaniach wykorzystano technologię ERP do zbadania wpływu braku snu na funkcje poznawcze, wczesne składniki, takie jak N1 i P2, nie były systematycznie badane, a wyniki są niespójne (Evans and Federmeier, 2007; Wiggins et al., 2018; Zhang et al., 2019). Istnieje kilka badań dotyczących zmiany składnika P2 podczas braku snu (Mograss et al., 2009). W związku z tym skutki braku snu dla wczesnych składników ERP, takich jak P2, nadal wymagają dalszych badań i badań.
w tym eksperymencie wykorzystaliśmy model 2-back do zaprojektowania zadań wymowy, pamięci przestrzennej i obiektowej oraz zbadaliśmy upośledzenie pamięci roboczej po 36 h TSD. W porównaniu z poprzednimi badaniami, które koncentrowały się tylko na wpływie braku snu na określony rodzaj informacji, takich jak wymowa pamięci roboczej lub specyficzne funkcje poznawcze, takie jak hamowanie odpowiedzi, rozważyliśmy zawartość modelu pamięci roboczej i kompleksowo przeanalizowaliśmy wpływ braku snu na pamięć roboczą.
jednak badanie ma pewne ograniczenia. Po pierwsze, użyliśmy tylko zadania 2-back i nie udało się porównać wydajności uczestników w zadaniach pamięci roboczej o różnych trudnościach. Dlatego istnieją ograniczenia w wyjaśnianiu i wnioskowaniu zmian w obciążeniu pracą. Po drugie, w badaniu wykorzystano wyłącznie ochotników płci męskiej,a wnioski należy ocenić, rozszerzając je na ochotniczki. Ze względu na ograniczoną liczbę uczestników stwierdziliśmy tylko, że amplituda fali N2 miała trend spadkowy, a opóźnienie fali P3 miało trend przedłużający się. Stabilne wyniki można uzyskać po zwiększeniu liczby uczestników. Po trzecie, połączenie naszej procedury z fMRI dla pamięci roboczej może ułatwić dalszą interpretację wyników. Wcześniejsze badania wykazały, że biorytmy okołodobowe wpływają na wydajność behawioralną i istnieją indywidualne różnice (Montplaisir, 1981; Lavie, 2001). Nie zarejestrowaliśmy danych EEG w tym samym czasie w tym eksperymencie, więc nie można całkowicie wykluczyć wpływu biorytmów okołodobowych na wyniki testu.
badania te wykazały, że zdolność pamięci roboczej była osłabiona po TSD i że uszkodzenie to nie było związane z zawartością bodźca w pamięci roboczej. Brak snu obniżył jakość informacji przechowywanych w pamięci, co może wystąpić wraz z degeneracyjnym procesem uwagi (Ratcliff and Van Dongen, 2018). Badanie to dostarcza dowodów elektrofizjologicznych na zrozumienie mechanizmu w zaburzonej pamięci roboczej po deprywacji snu. Konieczne jest zwrócenie uwagi na niekorzystne skutki upośledzenia pamięci roboczej spowodowane brakiem snu i zbadanie skutecznych interwencji w przypadku takich uszkodzeń.
Oświadczenie o dostępności danych
zbiory danych wygenerowane dla tego badania są dostępne na żądanie do odpowiedniego autora.
Oświadczenie o etyce
badania z udziałem ludzi zostały zweryfikowane i zatwierdzone przez czwarty Wojskowy Uniwersytet Medyczny Beihang University. Pacjenci / uczestnicy przedstawili pisemną świadomą zgodę na udział w tym badaniu.
wkład autora
Ys zaprojektował eksperymenty. ZP opracowało wyniki i napisało manuskrypt. CD i LZ analizowały i interpretowały dane. JT I YS przeprowadzili eksperymenty, zapoznali się z danymi i gwarancjami tego badania. YB, LZ i JT przyczyniły się do udziału w gromadzeniu danych i przeglądaniu literatury. Wszyscy wymienieni autorzy przeczytali i zatwierdzili ostateczny manuskrypt.
finansowanie
badania te były wspierane przez National Science Foundation of Winter Olympics Technology Plan Of China W ramach grantu nr 2019yff0301600 i HJ20191A020135.
konflikt interesów
autorzy oświadczają, że badania zostały przeprowadzone przy braku jakichkolwiek relacji handlowych lub finansowych, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Brak snu obniża hamowanie i zwiększa impulsywność do negatywnych bodźców. Zachowuj się. Brain Res. 217: 466. doi: 10.1016 / j.bbr.2010.09.020
PubMed Abstract | CrossRef Full Text | Google Scholar
Baddeley, A. (2000). Bufor epizodyczny: nowy składnik pamięci roboczej? Trendy Cogn. Sci. 4:417. doi: 10.1016/S1364-6613(00)01538-2
CrossRef Pełny tekst/Google Scholar
Bartel, P., Offermeier, W., Smith, F., and Becker, P. (2004). Uwaga i pamięć robocza u rezydentów anestezjologów po nocnej służbie: efekty grupowe i indywidualne. Occup. Environ. Med. 61, 167–170. doi: 10.1136 / oem.2002.006197
PubMed Abstract | CrossRef Full Text | Google Scholar
Casement, M. D., Broussard, J. L., Mullington, J. M., and Press, D. Z. (2006). Wkład snu w poprawę szybkości skanowania pamięci roboczej: badanie przedłużonego ograniczenia snu. Biol. Psychol. 72:212. doi: 10.1016 / j.biopsycho.2005.11.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Chee, M. W. L., and Chuah, L. Y. M. (2008). Funkcjonalne neuroobrazowanie wgląd w to, jak sen i brak snu wpływają na pamięć i poznanie. Curr. Opin. Neurol. 21, 417–423. doi: 10.1097 / WCO.0b013e3283052cf7
PubMed Abstract | CrossRef Full Text | Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S., and Chee, M. W. L. (2005). Dysocjacja regionów korowych modulowana zarówno przez obciążenie pamięci roboczej, jak i brak snu oraz przez brak snu. Neuroimage 25, 579-587. doi: 10.1016 / j.neuroimage.2004.11.029
PubMed Abstract | CrossRef Full Text | Google Scholar
Chuah, Y. M. L., Venkatraman, V., Dinges, D. F., and Chee, M. W. L. (2006). Neuronowa podstawa zmienności międzyosobniczej w skuteczności hamowania po deprywacji snu. J. Neurosci. 26, 7156–7162. doi: 10.1523 / jneurosci.0906-06.2006
CrossRef Pełny tekst | Google Scholar
Cote, K. A., Milner, C. E., Osip, S. L., Baker, M. L., and Cuthbert, B. P. (2008). Fizjologiczne pobudzenie i uwaga podczas Tygodnia ciągłego ograniczenia snu. Physiol. Zachowuj się. 95, 353–364. doi: 10.1016 / j.physbeh.2008.06.016
PubMed Abstract | CrossRef Full Text | Google Scholar
, Mondloch, C. J., Sergeeva, V., Taylor, M., and Semplonius, T. (2014). Wpływ całkowitego braku snu na behawioralne przetwarzanie nerwowe emocjonalnie ekspresyjnych twarzy. Exp. Brain Res. 232, 1429-1442. doi: 10.1007 / s00221-013-3780-1
PubMed Abstract | CrossRef Full Text/Google Scholar
Donchin, E., and Fabiani, M. (1991). Wykorzystanie potencjałów mózgu związanych z zdarzeniami w badaniu pamięci: czy p300 jest miarą odrębności zdarzeń? Psychol. Facul. Publiczne. 7, 58–77. doi: 10.1111 / sltb.12055
PubMed Abstract | CrossRef Full Text | Google Scholar
Drummond, S. P., Brown, G. G., Salamat, J. S., and Gillin, J. C. (2004). Rosnąca trudność zadania ułatwia mózgową reakcję kompensacyjną na całkowity brak snu. Sen 27, 445-451. doi: 10.1093/sleep / 27.3.445
CrossRef Full Text / Google Scholar
Wpływ całkowitego braku snu na mózgowe reakcje na funkcje poznawcze. Neuropsychofarmakologia 25, S68-S73. doi: 10.1016/S0893-133x(01)00325-6
CrossRef Full Text/Google Scholar
Pamięć, która jest w prawo i pamięć, która jest w lewo: potencjały związane z zdarzeniami ujawniają asymetrie półkuli w kodowaniu i zatrzymywaniu informacji werbalnych. Neuropsychologia 45, 1777-1790. doi: 10.1016 / j.neuropsychologia.2006.12.014
PubMed Abstract | CrossRef Full Text/Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M., and Höller, Y. (2007). Wizualny element p2 jest związany z blokowaniem fazowym theta. Neurosci. Lett. 426, 181–186. doi: 10.1016 / j.neulet.2007.08.062
PubMed Abstract | CrossRef Full Text | Google Scholar
Géraldine, R., Desgranges, B., Foret, J., and Eustache, F. (2005). Zależności między systemami pamięci a etapami snu. J. Sleep Res. 14, 123-140. doi: 10.1111 / j.1365-2869.2005.00450.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Gosselin, A., Koninck, J. D., and Campbell, K. B. (2005). Całkowity brak snu i nowe przetwarzanie: konsekwencje dla funkcjonowania płata czołowego. Clin. Neurofizjol. 116, 211–222. doi: 10.1016 / j.clinph.2004.07.033
PubMed Abstract | CrossRef Full Text | Google Scholar
Utrata snu upośledza krótkie i nowatorskie zadania językowe mające ostrość przedczołową. J. Śpiocha 7, 95-100 doi: 10.1046 / j.1365-2869.1998.00104.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J., and Meier, B. (2010). Jednoczesna Ważność zadania n-back jako miary pamięci roboczej. Pamięć 18, 394-412. doi: 10.1080/09658211003702171
PubMed Streszczenie | CrossRef Pełny tekst/Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., and Chamari, K. (2013). Wpływ częściowego braku snu na dobowe zmiany sprawności poznawczej u osób przeszkolonych. Proc. Soc. Zachowuj się. Sci. 82, 392–396. doi: 10.1016 / j.sbspro.2013.06.281
CrossRef Full Text | Google Scholar
Brak snu wpływa na niektóre, ale nie wszystkie procesy nadzoru uwagi. Psychol. Sci. 14, 473–479. doi: 10.1111/1467-9280.02456
PubMed Streszczenie | CrossRef Pełny tekst/Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L., and Lei, Y. (2015). Recovery sleep odwraca upośledzone zahamowanie odpowiedzi z powodu ograniczenia snu: dowody z badania potencjałów związanych z zdarzeniami wzrokowymi. PLoS One 10:e0142361 doi: 10.1371 / dziennik.pone.0142361
PubMed Abstract | CrossRef Full Text | Google Scholar
Funkcja płata czołowego, utrata snu i fragmentaryczny sen. Sleep Med. Obj. 5: 475. doi: 10.1053 / smrv.2001.0203
PubMed Abstract | CrossRef Full Text | Google Scholar
Koslowsky, M., and Babkoff, H. (1992). Metaanaliza związku między całkowitym brakiem snu a wydajnością. Chronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
PubMed Streszczenie | CrossRef Pełny tekst/Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A., and Hansenne, M. (2014). Nadużywanie alkoholu i Komponenty erp w zadaniach go / no-go z wykorzystaniem bodźców związanych z alkoholem: wpływ unikania alkoholu. Int. J. Psychofizjol. 94, 92–99. doi: 10.1016 / j.ijpsycho.2014.08.001
PubMed Streszczenie | CrossRef Pełny tekst | Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F., and Huster, R. J. (2019). Deprywacja snu w różny sposób wpływa na subkomponenty kontroli poznawczej. Sennik 42: zsz016 doi: 10.1093/sleep/zsz016
PubMed Abstract | CrossRef Full Text/Google Scholar
Sen-Budzenie jako rytm biologiczny. Annu. Ks. Psychol. 52, 277–303. doi: 10.1146 / annurev.psych.52.1.277
CrossRef Pełny tekst/Google Scholar
Lei, Y., Shao, Y., Wang, L., Zhai, T., and Zou, F. (2015). Wielkoskalowe sprzężenie sieci mózgowej przewiduje całkowity wpływ braku snu na zdolności poznawcze. PLoS One 10: e0133959. doi: 10.1371 / dziennik.pone.0133959
PubMed Streszczenie | CrossRef Pełny tekst/Google Scholar
Li, C., Huang, D., Qi, J., Chang, H., and Meng, Q. (2016). Deficyty pamięci krótkotrwałej korelują ze zmianami łączności czynnościowej hipokampa-talami po ostrym ograniczeniu snu. Obrazowanie Mózgu. 11, 954–963. doi: 10.1007 / s11682-016-9570-1
PubMed Streszczenie | CrossRef Pełny tekst/Google Scholar
Ma, N., Dinges, D. F., Basner, M., and Rao, H. (2014). Jak ostra całkowita utrata snu wpływa na mózg uczestniczący: metaanaliza badań neuroobrazowania. Sen 38, 233. doi: 10.5665 / sleep.4404
PubMed Abstract | CrossRef Full Text | Google Scholar
Miyake, A., and Shah, P. (1999). Modele pamięci roboczej. Mechanizmy aktywnego utrzymania i kontroli wykonawczej. Cambridge: Cambridge University Press.
Google Scholar
Mograss, M. A., Guillem, F., Brazzini-Poisson, V., and Godbout, R. (2009). Wpływ całkowitego braku snu na procesy rozpoznawania pamięci: badanie potencjału związanego z zdarzeniami. Neurobiol. Ucz się. Mem. 91, 343–352. doi: 10.1016 / j.nlm.2009.01.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Montplaisir, J. (1981). Depresja i rytm biologiczny: terapeutyczne skutki braku snu. L ’ Unión Médicale du Canada 110, 272-276. doi: 10.1620 / tjem.133.481
PubMed Abstract | CrossRef Full Text | Google Scholar
A., and Becker, C. E. (1992). Potencjał związany ze zdarzeniami p300. skutki braku snu. J. Occupat. Med. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F., And Hobson, J. A. (2002). Kora przedczołowa we śnie. Trendy Cogn. Sci. 6, 475–481. doi: 10.1016/S1364-6613(02)01992-7
CrossRef Pełny tekst/Google Scholar
Ning, M., Dinges, D. F., Mathias, B., and Hengyi, R. (2014). Jak ostra całkowita utrata snu wpływa na mózg uczestniczący: metaanaliza badań neuroobrazowania. Senna 38, 233-240 doi: 10.5665 / sleep.4404
PubMed Abstract | CrossRef Full Text | Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, A. R., and Bullmore, E. (2005). N-Back working memory paradigm: a meta-analysis of normative functional neuroimaging studies. Hum. MAPP Mózgu. 25, 46–59. doi: 10.1002 / hbm.20131
PubMed Streszczenie | CrossRef Pełny tekst | Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S., and Kumar, S. (2010). Elektrofizjologiczne korelaty funkcji poznawczych poprawiają się wraz z drzemką podczas braku snu. Eur. J. Appl. Physiol. 108, 549–556. doi: 10.1007 / s00421-009-1222-3
PubMed Streszczenie | CrossRef Pełny tekst/Google Scholar
Ratcliff, R., and Van Dongen, H. (2018). Wpływ braku snu na pozycję i pamięć asocjacyjną. J. Exp. Psychol. 44, 193–208. doi: 10.1037/xlm0000452
PubMed Abstract | CrossRef Full Text/Google Scholar
Zmiana pracy, cykle snu są na drodze do stania się kolejnym problemem zdrowia publicznego. JAMA J. am. Med. Assoc. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef Pełny tekst/Google Scholar
Saito, H., Yamazaki, H., Matsuoka, H., Matsumoto, K., and Sato, M. (2001). Wizualny potencjał związany ze zdarzeniami w łagodnym otępieniu typu Alzheimera. Psychiatria Clin. Neurosci. 55, 365–371. doi: 10.1046 / j. 1440-1819. 2001. 00876.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Schacht, A., Werheid, K., and Sommer, W. (2008). Ocena piękna twarzy jest szybka, ale nie obowiązkowa. Cogn. Afekt. Zachowuj się. Neurosci. 8, 132–142. doi: 10.3758 / cabn.8.2.132
PubMed Abstract | CrossRef Full Text | Google Scholar
Scott, J. P. R., Mcnaughton, L. R., and Polman, R. C. J. (2006). Wpływ braku snu i ćwiczeń na funkcje poznawcze, motoryczne i nastrój. Physiol. Zachowuj się. 87:408. doi: 10.1016 / j.physbeh.2005.11.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Smallwood, J., Brown, K.(2011) Współpraca między siecią trybu domyślnego a siecią czołowo-ciemieniową w produkcji wewnętrznego pociągu myśli. Brain Res. 1428, 60-70. doi: 10.1016 / j.brainres.2011.03.072
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, M. E., Mcevoy, L. K., and Gevins, A. (2002). Wpływ umiarkowanej utraty snu na sygnały neurofizjologiczne podczas wykonywania zadań z pamięcią roboczą. Sen 25, 784-794. doi: 10.1093/sleep / 25.7.56
CrossRef Full Text | Google Scholar
Neuronowe podstawy czujności i zaburzeń sprawności poznawczej w czasie senności ii. wpływ 48 i 72 h braku snu na przebudzenie aktywności Regionalnego mózgu człowieka. Thalam. Relat. Syst. 2, 199–229. doi: 10.1016/S1472-9288(03)00020-7
CrossRef Full Text/Google Scholar
Van Dongen, H. (2004). Łączny koszt dodatkowego czuwania: wpływ doseresponse na funkcje neurobehawioralne i fizjologię snu z przewlekłego braku snu i całkowitego braku snu. Sen 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef Pełny tekst / Google Scholar
Verweij, I. M., Romeijn, N., Smit, D. J., and Giovanni, P. (2014). Brak snu prowadzi do utraty funkcjonalnej łączności w przednich obszarach mózgu. BMC Neurosci. 15:88. doi: 10.1186/1471-2202-15-88
PubMed Abstract | CrossRef Full Text/Google Scholar
Wen, X., Liu, Y., Yao, L., and Ding, M. (2013). Odgórna Regulacja domyślnej aktywności trybu w przestrzennej uwagi wzrokowej. J. Neurosci. 33, 6444–6453. doi: 10.1523 / JNEUROSCI.4939-12. 2013
CrossRef Full Text | Google Scholar
24-godzinna deprywacja snu upośledza wczesną uważną modulację przetwarzania neuronowego: badanie potencjału mózgu związane z wydarzeniami. Neurosci. Lett. 677, 32–36. doi: 10.1016 / j.neulet.2018.04.022
PubMed Streszczenie | CrossRef Pełny tekst | Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y., and Zhou, Q. (2019). Zmniejszona wymiana informacji w pamięci roboczej po braku snu: dowody z badania potencjału związanego z wydarzeniami. Przód. Neurosci. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef Pełny tekst | Google Scholar
Zhang, Q., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z., et al. (2014). Badanie erp polegające na wzrokowym hamowaniu impulsów po drzemce wywołanej zaleplonem po deprywacji snu. PLoS One 9: e95653. doi: 10.1371 / dziennik.pone.0095653
PubMed Abstract | CrossRef Full Text | Google Scholar
Leave a Reply