grenzen in de neurowetenschappen
Inleiding
naarmate de maatschappij vordert en het werkritme verandert, lijdt een toenemend aantal mensen aan slaaptekort. Slaaptekort schaadt niet alleen de lichamelijke en geestelijke gezondheid van het individu, maar heeft ook ernstige gevolgen voor de werkprestaties, waardoor werkfouten en zelfs ongevallen ontstaan. Daarom is het begrijpen van het mechanisme van slaaptekort dat cognitieve functie beà nvloedt van groot belang voor het effectief verhinderen van de gevolgen van slaaptekort.eerdere studies hebben aangetoond dat slaaptekort een reeks veranderingen kan veroorzaken in iemands humeur, cognitief vermogen, werkprestaties en immuunfunctie (Choo et al., 2005). Het gebrek aan slaap verstoort de lichaamsomloop en beïnvloedt de cognitieve en emotionele vermogens van individuen (Raymond, 1988). Verschillende studies hebben aangetoond dat slaaptekort de remming van de respons belemmert (Harrison and Horne, 1998; Muzur et al., 2002; Jennings et al., 2003). Bijvoorbeeld, na 36 uur slaaptekort, verminderde het vermogen van het individu om negatieve stimuli te onderdrukken (Chuah et al., 2006). Neuroimaging studies hebben gesuggereerd dat slaapdeprivatie het lage niveau van visuele verwerkingscapaciteit van een individu vermindert (Anderson and platen, 2011; Ning et al., 2014). Bovendien schaadt slaaptekort de hippocampus en kan het geheugen beïnvloeden door synaptische plasticiteit te vernietigen (Cote et al., 2014). Thomas (2003) heeft aangegeven dat gebrek aan slaap verminderde cerebrale bloedstroom en stofwisseling in de thalamus, prefrontale cortex, en pariëtale cortex (Géraldine et al., 2005). Jarraya en collega ‘ s vonden dat gedeeltelijke slaaptekort neuropsychologische functies zoals verbale instantgeheugen, aandacht en alertheid aanzienlijk beïnvloedde (Thomas, 2003). Verder hebben sommige studies aangetoond dat de cumulatieve effecten van gedeeltelijke slaaptekort de cognitieve functie en het cognitieve gedrag ernstig zouden kunnen aantasten (van Dongen, 2004; Scott et al., 2006; Jarraya et al., 2013).
werkgeheugen is een systeem dat gebruikt wordt om informatie op te slaan en te verwerken en dat een cognitieve functie is met beperkte capaciteit (Bartel et al., 2004). Bovendien kan de informatie die in het werkende geheugensysteem wordt opgeslagen van geheugen op korte termijn in geheugen op lange termijn door het opnieuw vertellen en andere geheugenmethoden worden veranderd. Werkgeheugen is de overgang tussen korte-termijn-en lange-termijngeheugensystemen, die zeer centraal staat in de menselijke berichtenverwerking (Miyake and Shah, 1999). Het biedt een tijdelijke opslagruimte en de middelen die nodig zijn om informatie te verwerken, zoals spraakbegrip, redeneren en leren. Het is aangetoond dat slaaptekort het werkgeheugen eerst beïnvloedt.
eerdere studies hebben gebruik gemaakt van het n-back werkgeheugenparadigma bij deelnemers die slaaptekort ondergingen en vonden dat gebrek aan slaap een afname van metabole activiteit in het regionale netwerk van de hersenen veroorzaakt, die voornamelijk informatieverwerking en reactieremming veroorzaakt (Baddeley, 2000; Zhang et al., 2019). Verminderd werkgeheugen na slaaptekort is gerelateerd aan de activering van het standaardnetwerk in taken (Chee and Chuah, 2008), die gerelateerd kan zijn aan de belangrijke rol van de thalamus in corticale alertheid. Bijvoorbeeld, slaaptekort verhoogde de verbinding tussen de hippocampus, thalamus, en Standaard netwerk, die vaak gepaard ging met hogere subjectieve slaperigheid en slechtere prestaties van werkgeheugen (Lei et al., 2015; Li et al., 2016). Studies over slaaptekort identi?Rd dat verhoogde latentie en verminderde amplitude van de P3 component geassocieerd werden met langdurige soberheid (Morris et al., 1992; Jones and Harrison, 2001; Panjwani et al., 2010). De afname van de P3-golf zou een vermindering van de aandacht van de deelnemers en een vermindering van het onderscheiden van doelprikkels kunnen weerspiegelen (Koslowsky en Babkoff, 1992).
echter, weinig studies hebben elektrofysiologische aanwijzingen opgeleverd voor een verminderd werkgeheugen na slaaptekort. De n-back taak wordt beschouwd als een gemeenschappelijke methode om werkgeheugen te beoordelen (Owen et al., 2005; Jaeggi et al., 2010). Zhang et al. ontworpen een twee-back uitspraak werkgeheugen taak om de verminderde boodschap alternatief van werkgeheugen tijdens slaaptekort te verkennen, maar weinig studies hebben verschillende soorten werkgeheugen taken gebruikt in een enkel experiment. In deze studie ontwierpen we verschillende soorten werkgeheugentaken (uitspraak werkgeheugen, ruimtelijk werkgeheugen en objectgeheugen) om de verslechtering van cognitieve functie door TSD en geregistreerde EEG-gegevens van deelnemers op 2 tijdstippen (baseline en 36 H-TSD) te onderzoeken. Alle taken hebben een 2-back paradigma aangenomen. Deze studie evalueerde de veranderingen in de N2-P3 golf met betrekking tot werkgeheugen tijdens TSD en analyseerde de tijdelijke kenmerken van de effecten van slaapgebrek op werkgeheugen. Onze bevindingen leveren experimenteel bewijs voor de effecten van slaaptekort op cognitieve functies.
materialen en methoden
deelnemers
zestien jonge, gezonde, rechtshandig mannelijke studenten namen deel aan deze studie. We rekruteerden deelnemers door te adverteren op de campus. De deelnemers hadden allemaal goede slaapgewoonten (PSQI<5). Alle deelnemers waren tussen 21 en 28 jaar oud met een gemiddelde leeftijd van 23 jaar, en geen van de deelnemers had een psychische of lichamelijke ziekte. Alle deelnemers hadden normaal of gecorrigeerd zicht boven 1,0 en intelligentiescores >110 op de Raven-Test. Vóór het experiment legde de onderzoeker de procedure uit en wees de deelnemers erop dat ze vertrouwd waren met de methode en procedure. In de 2 weken voor het experiment sliepen de deelnemers regelmatig 7-9 uur per dag, zonder te roken, koffie te drinken, alcohol te drinken of medicijnen te consumeren gedurende 2 dagen voor het experiment. Vóór het experiment hebben alle deelnemers schriftelijke geïnformeerde toestemming gegeven. De experimentele regeling werd goedgekeurd door de Ethische Commissie van de vierde Militaire Medische Universiteit en Beihang University.
experimenteel ontwerp
drie typen werkgeheugentaken werden aan alle deelnemers gepresenteerd. Het waren twee-back uitspraak werkgeheugen taak( zie Figuur 1), twee-back ruimtelijke werkgeheugen taak (zie Figuur 2), en twee-back object werkgeheugen taak (zie Figuur 3). De stimulerende materialen van de taken waren 15 hoofdletterongevoelige Engelse letters die met uitzondering van degenen met soortgelijke letters, zoals L/l, M/m; kleine zwarte vierkantjes; en 12 geometrische figuren, respectievelijk. Alle materialen werden getoond in zwarte kleur op een witte achtergrond, met een geschatte visuele hoek van 1,5° × 1,5° (breedte: 2,0 cm, hoogte: 2,0 cm) subtiel. 122 trails werden opgenomen in elke taak en, in elk trail, de doel stimulus werd gepresenteerd voor 400 ms twee trails na de presentatie van objectieve stimulus, met de 1600 ms stimulus begin asynchrony time (SOA) die werd gemarkeerd door een witte “+.”De deelnemers werd gevraagd om op de linker muisknop te klikken wanneer het doel en de objectieve stimulus hetzelfde waren (“matching”), terwijl ze op de rechter muisknop klikten wanneer ze dat niet waren (“mismatching”). De matching or not voorwaarde werden gepresenteerd in een pseudorandom orde met een 1: 1 verhouding.
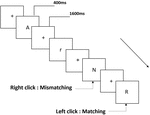
figuur 1. Schematisch diagram van de uitspraak werkgeheugen taak.
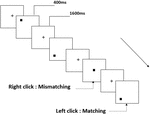
Figuur 2. Schematisch schema van de ruimtelijke werkgeheugent taak.
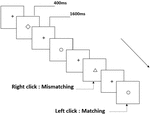
Figuur 3. Schematisch diagram van de taak van het werkgeheugen van het object.
experimentele Procedures
vóór het experiment werden de deelnemers geïnstrueerd over de experimentele taak. Zij werden geà nformeerd om de drie soorten werkgeheugentaken uit te oefenen totdat een nauwkeurigheid van 90% werd bereikt. De deelnemers bezochten het laboratorium de dag voor het experiment en sliepen die nacht in het laboratorium. De twee partnerdeelnemers voerden de experimenten tegelijkertijd uit. Drie soorten werkgeheugentaken werden uitgevoerd om 7: 30 tot 8: 30 de volgende ochtend met gelijktijdige elektro-encefalogram (EEG) opname (baseline). De tweede EEG-opname (36 H-TSD) werd uitgevoerd na een periode van 36 h waarin de deelnemers niet mochten slapen. Gedurende de gehele proeftijd waren centrale remming en stimulerende middelen verboden. De deelnemers werden begeleid, geobserveerd en herinnerd door het verplegend personeel om hen tijdens de TSD-sessie wakker te houden.
EEG opnames
een continue scalp EEG werd opgenomen met behulp van elektrodedoppen geplaatst op 64 locaties met behulp van het 10-20 systeem met een SynAmps2 versterker. De bilaterale mastoïden (A1 en A2) werden gebruikt als referentie, en het voorhoofd werd gebruikt als grond. EEG ‘ s werden geregistreerd bij 1000 Hz en de impedantie van alle kanalen bleef onder de 5 kΩ. Vier extra elektroden werden boven en onder de rechter en linker ogen geplaatst om een bipolair verticaal en horizontaal elektrooculogram op te nemen.
gegevensanalyse van gedragsexperimenten
vanwege technische fouten werden twee gevallen verwijderd, terwijl andere 14 gevallen werden opgenomen in de volgende statistische analyse. Gedragsgegevens omvatten de gemiddelde reactietijd, de juiste snelheid en het juiste aantal per tijdseenheid. Gedragsgegevens in baseline en 36 h-TSD toestanden werden geregistreerd voor analyse. De analyses werden uitgevoerd door IBM SPSS (V22.2), waar de herhaalde metingen analyse van variantie (ANOVA) methode met Greenhouse-Geisser werd Bonferroni post-hoc analyse werd gelanceerd. De statistische resultaten werden gepresenteerd als gemiddelde en standaardafwijking (SD).
EEG-gegevensanalyse
Scan 4.3 programma werd gebruikt om de EEG-gegevens te analyseren, waarbij de EEG-artefacten van de oogbeweging werden gecorrigeerd door middel van oculaire artefactreductiemethode. Tijdperken variërend van -100 tot 800 ms van de continue EEG-gegevens werden geëxtraheerd en gefilterd door een bandpasfilter van 0,5 tot 30 Hz met de frequentiehelling van 24 dB/oct. De onderzoeken waarbij de spanning meer dan ± 100 µV bedroeg, werden afgewezen en de uitgangswaarde werd gecorrigeerd tot een gemiddelde amplitude van 100 ms. de EEG-componenten werden gemiddeld en berekend met alleen de gecorrigeerde responsen. De ERP-componenten P2 (100-250 ms), N2 (150-350 ms) en P3 (250-450 ms) van de stimulusstudies werden geïdentificeerd en gekwantificeerd. De grootgemiddelde piekamplitudes en latenties van de N2-en P3-componenten werden afzonderlijk berekend bij F3, Fz, F4, C3, Cz, C4, P3, Pz en P4, en de P2-component werd berekend bij F3, FZ, F4, C3, Cz en C4 (Casement et al., 2006; Verweij et al., 2014).
herhaalde metingen ANOVA werd gebruikt voor alle ERP-resultaten. De belangrijkste effecten en de interacties tussen slaaptoestanden (baseline en 36 H-TSD), taken (uitspraak werkgeheugen, ruimtelijk werkgeheugen en objectgeheugen), regio ’s (frontaal, centraal en pariëtaal; de P2 component werd alleen geanalyseerd op de frontale en centrale regio’ s), en sites (Links, Midden en rechts) werden statistisch geanalyseerd met behulp van herhaalde metingen ANOVA, waaronder Kasgeisser correcties voor niet-bolvormigheid en Bonferroni post-hoc testen.
resultaten
Gedragsprestaties
de resultaten van de gedragsexperimenten worden weergegeven in Tabel 1. De gemiddelde reactietijd was langer in de 36 H-TSD-toestand dan bij baseline, met een trend om toe te nemen, maar zonder significante verschillen. ANOVA toonde aan dat het juiste percentage van de taak significant verschilde tussen de baseline en 36 H-TSD . Het juiste aantal per tijdseenheid toonde een significant hoofdeffect van de tijd tijdens 36 H-TSD .

Tabel 1. Prestatiegegevens (gemiddelde ± SD) van de 2-back-taak bij baseline en na 36 H-TSD.
Amplitude
vergeleken met de uitgangswaarde werd een significante afname in de amplitude van P3 waargenomen en een significante toename in de amplitude van P2 na TSD. Hoewel de N2-amplitude na 36 uur TSD daalde, bereikte het verschil geen statistische significantie (Tabel 2).

Tabel 2. Grootgemiddelde piekamplitude van de P2 -, N2-en P3-componenten in de juiste responsconditie over meerdere elektrodeplaatsen bij baseline en na 36 H-TSD.
significante belangrijke effecten van regio ‘ s en locaties op de P2-amplitude werden gevonden onder de TSD-voorwaarde. Tijdens TSD verscheen de maximale amplitude van P2 in het frontale gebied (Figuur 4). Bovendien waren de verschillen in P2 amplitudes in verschillende gebieden (frontaal vs.centraal) groter in de bilaterale elektroden (links: P = 0,001; rechts: P = 0.000) dan in de middelste elektroden (Figuur 4). Een significant belangrijk effect van het gebied en de plaats op N2 toonde aan dat de N2 amplitude negatiever was in het frontale dan in het centrale gebied (P = 0,008, Figuur 4) en kleiner was aan de rechterkant dan aan de linkerkant (P = 0,011, figuren 5A2,B2). Een belangrijk effect van site op de P3 amplitude werd waargenomen . De amplitude van P3 was positiever in het midden dan aan de linkerkant (P = 0,009, Figuur 4). Een significant interactieeffect tussen tijd en gebied werd waargenomen voor de P3-amplitude . Tijdens TSD was de reductie in P3 amplitude significanter in de frontale en centrale gebieden dan in het pariëtale gebied (P = 0,005; P = 0,003) (figuur 5C3). Geen andere belangrijke effecten of interactieeffecten bereikten statistische significantie.
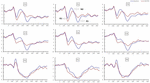
Figuur 4. ERP-amplitude bij baseline en 36 H-TSD voor de juiste responsconditie voor de werkgeheugentaak. De kanalen worden als volgt van links naar rechts en van boven naar beneden geordend: F3, Fz, F4, C3, Cz, C4, P3, Pz en P4. Vergeleken met de baseline waren de latenties van de N2-P3 componenten verlengd en de amplitudes van N2-P3 waren verlaagd na 36 H-TSD.
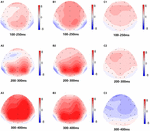
Figuur 5. Topografische kaart van de juiste respons in het werkgeheugen taak in verschillende slaapomstandigheden (A1-C3). (A1) P2, 100-250 ms, bij baseline. (A2) N2, 200-300 ms, bij baseline. (A3) P3, 300-400 ms, bij baseline. (B1) P2, 100-250 ms, bij 36 H-TSD. (B2) N2, 200-300 ms, voor 36 H-TSD. (B3) P3, 300-400 ms, voor 36 H-TSD. (C1) P2, 100-250 ms, 36 H-TSD met afgetrokken baseline. (C2) N2, 200-300 ms, 36 H-TSD met aftrek van de uitgangswaarde. (C3) P3, 300-400 ms, 36 H-TSD met aftrek van de uitgangswaarde.
latentie
De latenties van N2 en P2 waren significant verlengd na TSD. Hoewel de P3-latentie verlengd was na 36 uur TSD, bereikte het verschil geen statistische significantie (Tabel 3).

Tabel 3. Grand-average peak latency van de P2, N2 en P3 componenten in de juiste responsconditie over meerdere elektrodeplaatsen bij baseline en na 36 H-TSD.
het significante belangrijkste effect van regio op N2 en P3 toonde aan dat de latentie van de N2-P3 componenten korter was in het pariëtale gebied dan in het frontale gebied (P = 0,002; P = 0.000) en het centrale gebied (P = 0.000; P = 0.000) (Figuur 4). De latentie van de P3 golf was significant langer aan de linkerkant dan aan de rechterkant (Figuur 4).
geen andere belangrijke effecten of interactieeffecten bereikten statistische significantie.
de N2, P2, en P3 amplitudes en latenties die werden opgewekt op de negen elektrode locaties worden weergegeven in Figuur 4. De topografische kaart van de juiste respons in het werkgeheugen taak in verschillende slaapomstandigheden (baseline, 36 H-TSD en het verschil tussen de twee voorwaarden) is weergegeven in Figuur 5.
discussie
in deze studie rapporteerden we de invloeden van 36 h slaapdeprivatie op het werkgeheugen, waarbij gedragsgegevens in twee slaapstaten (baseline en 36 H-TSD) werden gecombineerd met gelijktijdige EEG-opnames. De analyse van de resultaten gaf aan dat de veranderingen in de gedragsgegevens in overeenstemming met het werkgeheugen na 36 uur TSD verslechterden: een toename van de gemiddelde reactietijd van de cognitieve taken en een afname van de nauwkeurigheid.
slaaptekort verstoorde de individuele controle over de aandachtsmiddelen. Hoewel individuen probeerden om waakzaamheid en werkprestaties, met inbegrip van de reactietijd en correcte snelheid te handhaven, tijdens slaaptekort, werd de capaciteit van de informatieverwerking van hun werkgeheugen nog beïnvloed wegens de daling van de snelheid van het verwerken van informatie (Casement et al., 2006; Wiggins et al., 2018). In deze studie werden de N2-en P3-golven gerelateerd aan het werkgeheugen gemeten om een toename van de latentie en een afname van de amplitude na slaaptekort te laten zien in vergelijking met de uitgangswaarden. Studies hebben aangetoond dat slaaptekort leidt tot een continue afname van de aandacht, en het fenomeen van verminderde P3 amplitude wijst erop dat de top-down controle van kennis van individuen geleidelijk instort. Slaaptekort heeft een meer negatief effect op cognitieve functies, vooral die welke afhankelijk zijn van mentale of cognities (Kusztor et al., 2019).
de P3-component weerspiegelt de inzet van aandachtsmiddelen, en de latency van P3 wordt algemeen gezien als het tijdvenster voor stimulus categorisatie en evaluatie. De afname van de P3-golfamplitude bevestigde ook dat de besluitvorming in de matching respons na TSD tot op zekere hoogte was beschadigd (Gosselin et al., 2005). De Studies hebben gesuggereerd dat slaapgebrek het stadium van de informatieverwerking van werkgeheugen kan beà nvloeden. In deze studie ondersteunden de prestatie-indicatoren ook de conclusie dat de responstijd op de doelprikkel was verhoogd en dat de latentie van de P3-golf was verlengd (Cote et al., 2008). Er werd gespeculeerd dat het effect van slaaptekort op P3-componenten ook zou kunnen plaatsvinden vanwege het niet reageren op informatieverandering, wat in overeenstemming is met eerdere conclusies dat de P3-componenten gerelateerd zijn aan het bijwerken van werkgeheugeninhoud (Donchin en Fabiani, 1991).
eerdere studies hebben de N2-component beschouwd als een elektrofysiologische index die het vermogen van het individu om de respons te onderdrukken weergeeft (Kreusch et al., 2014). Na slaaptekort geeft de verlengde latentie van de Nogo-N2 component aan dat het vermogen van het individu om de respons te onderdrukken verminderd is (Jin et al., 2015). De verminderde amplitude en verlengde latentie van de N2-component met betrekking tot uitspraak werkgeheugen na slaaptekort onthult dat slaaptekort de informatieverwerking van uitspraak werkgeheugen schaadt (Zhang et al., 2019). De N2 component wordt over het algemeen verondersteld om de selectieve aandacht van de hersenen en de verwerking van emotionele stimuli of signalen weer te geven (Schacht et al., 2008) en is een endogene component gerelateerd aan de mentale toestand van een individu, aandacht, en de mate van aandacht. In deze studie vonden we dat de latentie van de N2 component aanzienlijk verlengd was, maar de amplitude vertoonde slechts een neerwaartse trend. Volgens eerdere studies weerspiegelde de verlengde N2-latentie een toename van de responstijd na slaapbeperking (Zhang et al., 2014). De bevinding dat N2-amplitude niet significant werd gewijzigd, kan echter te wijten zijn aan cerebrale compenserende responsen (Drummond and Brown, 2001). In het geval van beperkte cognitieve middelen was er een compensatiemechanisme om een verminderde cognitieve functie te herstellen (Jin et al., 2015).
volgens de topografie van de hoofdhuid zijn de veranderingen in de N2-P3 componenten gerelateerd aan slaaptekort duidelijker in het frontale gebied. Frontoparietale controle (FPC) speelt een belangrijke rol in cognitieve controle. Studies hebben aangetoond dat FPC kan omzeilen top-down cognitieve controle, waardoor individuen om zich te concentreren op informatie met betrekking tot het doel, terwijl het onderdrukken van informatie die niet is gerelateerd aan het doel (Smallwood et al., 2011; Wen et al., 2013). FPC is belangrijk voor het bewaren en verwerken van informatie in het werkgeheugen, en de mate van activering van FPC na slaaptekort was verminderd in vergelijking met een normale slaapgroep (Ma et al., 2014). Hoewel de EEG-resultaten de veranderingen in specifieke hersengebieden niet in detail weerspiegelden, weerspiegelde het intuïtief het effect van TSD op de retentie en verwerking van werkgeheugeninformatie.
hoewel het exacte cognitieve proces dat de P2 component ten grondslag ligt nog steeds breed wordt besproken, als een brede definitie, weerspiegelt de P2 component het proces van aandacht en visuele verwerking en wordt algemeen beschouwd als gerelateerd aan selectieve aandacht en werkgeheugen, als gevolg van het vroege oordeel van het perceptuele proces (Saito et al., 2001). In deze studie vonden we een significante toename van de amplitude van de P2-golf na slaaptekort. Studies hebben gemeld dat P2 golven die een deel van de vroege cognitieve matching systeem voor berichtverwerking zou kunnen zijn en zintuiglijke input aan opgeslagen geheugen kunnen vergelijken (Freunberger et al., 2007) gevoelig zijn voor veranderingen in de aandacht van de missie en het werkgeheugen eisen (Smith et al., 2002). Functionele compensatie is een van de unieke functies van de menselijke hersenen en een belangrijke factor voor het handhaven van cognitieve functie. Overmatige activering van de dorsolaterale prefrontale cortex (DLPFC) na slaaptekort geeft aan dat, als de hersenbronnen afnemen, de DLPFC een compenserende functie lijkt te hebben (Drummond et al., 2004; Choo et al., 2005). Daarom speculeren we dat de significante toename van P2 amplitude waargenomen in deze studie te wijten kan zijn aan functionele compensatie waarbij individuen lijken te handhaven normale cognitieve functie na slaaptekort. Hoewel een groot aantal studies ERP-technologie hebben gebruikt om het effect van slaaptekort op cognitieve functies te onderzoeken, zijn vroege componenten zoals N1 en P2 niet systematisch onderzocht, en de resultaten zijn inconsistent (Evans and Federmeier, 2007; Wiggins et al., 2018; Zhang et al., 2019). Er zijn weinig onderzoeken onderzoeken de verandering van P2 component tijdens slaaptekort (Mograss et al., 2009). Daarom moeten de effecten van slaaptekort op vroege componenten van ERP, zoals P2, nog verder worden bestudeerd en onderzocht.
in dit experiment gebruikten we het 2-back model om uitspraak, ruimtelijke en object werkgeheugentaken te ontwerpen en onderzochten we de beperking van werkgeheugen na 36 uur TSD. Vergeleken met eerdere studies die alleen gericht op de effecten van slaaptekort op een specifiek type informatie, zoals uitspraak werkgeheugen, of specifieke cognitieve functie, zoals reactie remming, hebben we de inhoud van het werkgeheugenmodel overwogen en uitgebreid de effecten van slaaptekort op het werkgeheugen geanalyseerd.
het onderzoek heeft echter enkele beperkingen. Ten eerste hebben we alleen de 2-back-taak gebruikt en zijn we er niet in geslaagd om de prestaties van de deelnemers in werkgeheugentaken van verschillende moeilijkheden te vergelijken. Daarom zijn er beperkingen in het verklaren en afleiden van veranderingen in werklast. Ten tweede werden in het onderzoek alleen mannelijke vrijwilligers gebruikt en de conclusies moeten worden beoordeeld wanneer ze worden uitgebreid tot vrouwelijke vrijwilligers. Door het beperkte aantal deelnemers, vonden we alleen dat de amplitude van de N2 golf een neerwaartse trend had en dat de P3 Golf latentie een verlengde trend had. Stabiele resultaten kunnen worden verkregen na het verhogen van het aantal deelnemers. Ten derde kan het combineren van onze procedure met fMRI voor werkgeheugen verdere interpretatie van de resultaten vergemakkelijken. Eerdere studies hebben aangetoond dat circadiaanse bioritmen de gedragsprestaties beïnvloeden, en er zijn individuele verschillen (Montplaisir, 1981; Lavie, 2001). We hebben de EEG-gegevens niet op hetzelfde tijdstip in dit experiment vastgelegd, dus de invloed van circadiaanse bioritmen op de testresultaten kan niet volledig worden uitgesloten.
Dit onderzoek toonde aan dat de capaciteit van het werkgeheugen werd verminderd na TSD en dat deze schade niet in verband werd gebracht met de stimulusinhoud van het werkgeheugen. Het gebrek aan slaap verminderde de kwaliteit van de in het geheugen opgeslagen informatie, die kan optreden bij het degeneratieve proces van aandacht (Ratcliff en van Dongen, 2018). Deze studie levert elektrofysiologisch bewijs voor het begrijpen van het mechanisme onder het verminderde werkgeheugen na slaaptekort. Het is noodzakelijk om aandacht te besteden aan de nadelige effecten van het werkgeheugen, veroorzaakt door slaaptekort, en om effectieve interventies voor dergelijke schade te onderzoeken.
Gegevensbeschikbaarheidsverklaring
de voor dit onderzoek gegenereerde datasets zijn op verzoek beschikbaar voor de overeenkomstige auteur.
Ethics Statement
de studies waarbij menselijke deelnemers betrokken waren, werden beoordeeld en goedgekeurd door de vierde Militaire Medische Universiteit van Beihang University. De patiënten / deelnemers hebben hun schriftelijke geïnformeerde toestemming gegeven om deel te nemen aan deze studie.
Auteursbijdragen
YS ontwierp de experimenten. ZP produceerde de resultaten en schreef het manuscript. CD en LZ analyseerden en interpreteerden de gegevens. JT en YS voerden de experimenten uit, maakten kennis met de gegevens en de garanten van deze studie. YB, LZ en JT hebben bijgedragen aan de deelname aan het verzamelen van gegevens en het herzien van de literatuur. Alle genoemde auteurs hebben het definitieve manuscript gelezen en goedgekeurd.
financiering
Dit onderzoek werd ondersteund door de National Science Foundation of Winter Olympics Technology Plan of China onder subsidie nrs. 2019YFF0301600 en HJ20191A020135.
belangenconflict
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Anderson, C., and Platten, C. R. (2011). Slaaptekort verlaagt remming en verbetert impulsiviteit tot negatieve stimuli. Behav. Brain Res. 217: 466. doi: 10.1016 / j.bbr.2010.09.020
PubMed Abstract / CrossRef Full Text / Google Scholar
Baddeley, A. (2000). De episodische buffer: een nieuw onderdeel van werkgeheugen? Trends Cogn. Sci. 4:417. doi: 10.1016/s1364-6613(00)01538-2
CrossRef Full Text/Google Scholar
Bartel, P., Offermeier, W., Smith, F., and Becker, P. (2004). Aandacht en werkgeheugen bij inwonende anesthesisten na nachtdienst: groepseffecten en individuele effecten. Occup. Environ. Med. 61, 167–170. doi: 10.1136 / oem.2002.006197
PubMed Abstract / CrossRef Full Text / Google Scholar
Casement, M. D., Broussard, J. L., Mullington, J. M., and Press, D. Z. (2006). De bijdrage van slaap aan verbeteringen in de snelheid van het scannen van het werkgeheugen: een studie van langdurige slaapbeperking. Biol. Psychol. 72:212. doi: 10.1016 / j.biopsycho.2005.11.002
PubMed Abstract / CrossRef Full Text / Google Scholar
Chee, M. W. L., and Chuah, L. Y. M. (2008). Functionele neuroimaging inzichten in hoe slaap en slaaptekort het geheugen en de cognitie beïnvloeden. Curr. Opin. Neurol. 21, 417–423. doi: 10.1097 / WCO.0b013e3283052cf7
PubMed Abstract / CrossRef Full Text / Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S., and Chee, M. W. L. (2005). Dissociatie van corticale regio ‘ s gemoduleerd door zowel werkgeheugenbelasting en slaaptekort en alleen door slaaptekort. Neuroimage 25, 579-587. doi: 10.1016 / j.neuroimage.2004.11.029
PubMed Abstract / CrossRef Full Text / Google Scholar
Chuah, Y. M. L., Venkatraman, V., Dinges, D. F., and Chee, M. W. L. (2006). De neurale basis van interindividuele variabiliteit in remmende efficiëntie na slaaptekort. J. Neurosci. 26, 7156–7162. doi: 10.1523 / jneurosci.0906-06. 2006
CrossRef Full Text / Google Scholar
Cote, K. A., Milner, C. E., Osip, S. L., Baker, M. L., and Cuthbert, B. P. (2008). Fysiologische opwinding en aandacht tijdens een week van continue slaapbeperking. Fysiol. Behav. 95, 353–364. doi: 10.1016 / j. physbeh.2008.06.016
PubMed Abstract | CrossRef Full Text/Google Scholar
Cote, K. A., Mondloch, C. J., Sergeeva, V., Taylor, M., and Semplonius, T. (2014). Impact van totale slaaptekort op gedragsneurale verwerking van emotioneel expressieve gezichten. Exp. Brain Res. 232, 1429-1442. doi: 10.1007 / s00221-013-3780-1
PubMed Abstract / CrossRef Full Text / Google Scholar
Donchin, E., and Fabiani, M. (1991). Het gebruik van event-related brain potentials in de studie van het geheugen: is p300 een maat voor event onderscheidend vermogen? Psychol. Facul. Openbaar. 7, 58–77. doi: 10.1111 / sltb.12055
PubMed Abstract / CrossRef Full Text / Google Scholar
Drummond, S. P., Brown, G. G., Salamat, J. S., and Gillin, J. C. (2004). Toenemende taak moeilijkheid vergemakkelijkt de cerebrale compenserende reactie op totale slaaptekort. Slaap 27, 445-451. doi: 10.1093/sleep/27.3.445
CrossRef Full Text / Google Scholar
Drummond, S. P. A., and Brown, G. G. (2001). De effecten van totale slaaptekort op cerebrale reacties op cognitieve prestaties. Neuropsychopharmacologie 25, S68-S73. doi: 10.1016/S0893-133X(01)00325-6
CrossRef Full Text/Google Scholar
Evans, K. M., and Federmeier, K. D. (2007). Het geheugen dat\”s rechts en het geheugen dat \” s links: event-gerelateerde potentialen onthullen hemisferische asymmetrieën in de codering en retentie van verbale informatie. Neuropsychologia 45, 1777-1790. doi: 10.1016 / j.neuropsychologia.2006.12.014
PubMed Abstract / CrossRef Full Text / Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M., and Höller, Y. (2007). Visuele P2 component is gerelateerd aan Theta fase-vergrendeling. Neurowetenschappen. Lett. 426, 181–186. doi: 10.1016 / j.neulet.2007.08.062
PubMed Abstract / CrossRef Full Text / Google Scholar
Géraldine, R., Desgranges, B., Foret, J., and Eustache, F. (2005). De relaties tussen geheugensystemen en slaapstadia. J. Sleep Res. 14, 123-140. doi: 10.1111 / j. 1365-2869.2005.00450.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Gosselin, A., Koninck, J. D., and Campbell, K. B. (2005). Totale slaaptekort en nieuwe verwerking: implicaties voor de werking van de frontale kwab. Clin. Neurofysiol. 116, 211–222. doi: 10.1016 / j.clinph.2004.07.033
PubMed Abstract / CrossRef Full Text / Google Scholar
Harrison, Y., and Horne, J. (1998). Slaapverlies schaadt korte en nieuwe taaltaken met een prefrontale focus. J. Sleep Res. 7, 95-100. doi: 10.1046 / j. 1365-2869. 1998. 00104.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J., and Meier, B. (2010). De gelijktijdige geldigheid van de n-back-taak als werkgeheugenmaat. Geheugen 18, 394-412. doi: 10.1080 / 09658211003702171
PubMed Abstract / CrossRef Full Text / Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., and Chamari, K. (2013). De impact van gedeeltelijke slaaptekort op de dagelijkse variaties van cognitieve prestaties bij getrainde proefpersonen. Proc. Soc. Behav. Sci. 82, 392–396. doi: 10.1016 / j. sbspro.2013.06.281
CrossRef Full Text / Google Scholar
Jennings, J. R., Monk, T. H., and Molen, M. W. V. D. (2003). Slaaptekort beïnvloedt sommige maar niet alle processen van toezicht. Psychol. Sci. 14, 473–479. doi: 10.1111 / 1467-9280. 02456
PubMed Abstract / CrossRef Full Text / Google Scholar
Jin, X., Ye, E., Qi, J., Wang, L. en Lei, Y. (2015). Herstelslaap keert verminderde remming van de respons als gevolg van slaapbeperking om: bewijs van een visueel event-gerelateerd potentiaalonderzoek. PLoS One 10:e0142361. doi: 10.1371 / journal.pone.0142361
PubMed Abstract / CrossRef Full Text / Google Scholar
Jones, K., and Harrison, Y. (2001). Frontale kwabfunctie, slaapverlies en gefragmenteerde slaap. Slaap Med. Openbaring 5: 475. doi: 10.1053 / smrv.2001.0203
PubMed Abstract / CrossRef Full Text / Google Scholar
Koslowsky, M., and Babkoff, H. (1992). Meta-analyse van de relatie tussen totale slaaptekort en prestaties. Chronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
PubMed Abstract / CrossRef Full Text / Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A., and Hansenne, M. (2014). Alcoholmisbruik en erp componenten in go / no-go taken met behulp van alcoholgerelateerde stimuli: impact van alcoholvermijding. Int. J. Psychophysiol. 94, 92–99. doi: 10.1016 / j. ijpsycho.2014.08.001
PubMed Abstract / CrossRef Full Text / Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F., and Huster, R. J. (2019). Slaaptekort heeft een verschillende invloed op subcomponenten van cognitieve controle. Slaap 42: zsz016. doi: 10.1093 / sleep / zsz016
PubMed Abstract / CrossRef Full Text / Google Scholar
Lavie, P. (2001). Slaap-wake als een biologisch ritme. Annu. Rev. Psychol. 52, 277–303. doi: 10.1146 / annurev.psych.52.1.277
CrossRef Full Text / Google Scholar
Lei, Y., Shao, Y., Wang, L., Zhai, T., en zou, F. (2015). Grootschalige netwerkkoppeling van hersenen voorspelt totale slaapdeprivatie-effecten op cognitieve capaciteit. PLoS One 10: e0133959. doi: 10.1371 / journal.pone.0133959
PubMed Abstract / CrossRef Full Text / Google Scholar
Li, C., Huang, D., Qi, J., Chang, H., and Meng, Q. (2016). Korte-termijn geheugen tekorten correleren met hippocampal-thalamische functionele connectiviteit veranderingen na acute slaap beperking. Brain Imaging Behav. 11, 954–963. doi: 10.1007 / s11682-016-9570-1
PubMed Abstract / CrossRef Full Text / Google Scholar
Ma, N., Dinges, D. F., Basner, M., and Rao, H. (2014). Hoe acuut totaal slaapverlies de aanwezige hersenen beïnvloedt: een meta-analyse van neuroimaging studies. Slaap 38, 233. doi: 10.5665 / slaap.4404
PubMed Abstract / CrossRef Full Text / Google Scholar
Miyake, A., and Shah, P. (1999). Modellen van werkgeheugen. Mechanismen van actief onderhoud en uitvoerende controle. Cambridge: Cambridge University Press.
Google Scholar
Mograss, M. A., Guillem, F., Brazzini-Poisson, V., and Godbout, R. (2009). The effects of total sleep deprivation on recognition memory processes: een studie van event-related potential. Neurobiol. Leren. Mem. 91, 343–352. doi: 10.1016 / j.nlm.2009.01.008
PubMed Abstract / CrossRef Full Text / Google Scholar
Montplaisir, J. (1981). Depressie en biologisch ritme: therapeutische effecten van slaaptekort. L ‘ Unión Médicale du Canada 110, 272-276. doi: 10.1620 / tjem.133.481
PubMed Abstract / CrossRef Full Text / Google Scholar
Morris, A. M., So, Y., Lee, K. A., Lash, A. A., and Becker, C. E. (1992). Het P300 event-gerelateerde potentieel. de effecten van slaaptekort. J. Occupat. Med. 34, 1143–1152.
Google Scholar
Muzur, A., Pace-Schott, E. F., and Hobson, J. A. (2002). De prefrontale cortex in slaap. Trends Cogn. Sci. 6, 475–481. doi: 10.1016/S1364-6613(02)01992-7
CrossRef Full Text/Google Scholar
Ning, M., Dinges, D. F., Mathias, B., and Hengyi, R. (2014). Hoe acuut totaal slaapverlies de aanwezige hersenen beïnvloedt: een meta-analyse van neuroimaging studies. Slaap 38, 233-240. doi: 10.5665 / slaap.4404
PubMed Abstract / CrossRef Full Text / Google Scholar
Owen, A. M., Mcmillan, K. M., Laird, A. R., and Bullmore, E. (2005). N-back werkgeheugenparadigma: een meta-analyse van normatieve functionele neuroimaging studies. Brom. Brain Mapp. 25, 46–59. doi: 10.1002 / hbm.20131
PubMed Abstract / CrossRef Full Text / Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, S., and Kumar, S. (2010). Elektrofysiologische correlaten van cognitie verbeteren met nap tijdens slaaptekort. Euro. J. Appl. Fysiol. 108, 549–556. doi: 10.1007 / s00421-009-1222-3
PubMed Abstract / CrossRef Full Text / Google Scholar
Ratcliff, R., and Van Dongen, H. (2018). De effecten van slaaptekort op item en associatief herkenningsgeheugen. J. Exp. Psychol. 44, 193–208. doi: 10.1037 / xlm0000452
PubMed Abstract / CrossRef Full Text / Google Scholar
Raymond, C. A. (1988). Verschuivende arbeid, slaapcycli zijn op weg om een ander volksgezondheidsvraagstuk te worden. JAMA J. Am. Med. Assoc. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef Full Text / Google Scholar
Saito, H., Yamazaki, H., Matsuoka, H., Matsumoto, K., and Sato, M. (2001). Visueel event-gerelateerd potentieel bij milde dementie van het alzheimer-type. Psychiatrie Clin. Neurowetenschappen. 55, 365–371. doi: 10.1046 / j. 1440-1819. 2001. 00876.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Schacht, A., Werheid, K., and Sommer, W. (2008). De beoordeling van gezichtschoonheid is snel, maar niet verplicht. Cogn. Invloed. Behav. Neurowetenschappen. 8, 132–142. doi: 10.3758 / cabn.8.2.132
PubMed Abstract / CrossRef Full Text / Google Scholar
Scott, J. P. R., Mcnaughton, L. R., and Polman, R. C. J. (2006). Effecten van slaaptekort en lichaamsbeweging op cognitieve, motorische prestaties en stemming. Fysiol. Behav. 87:408. doi: 10.1016 / j. physbeh.2005.11.009
PubMed Abstract / CrossRef Full Text / Google Scholar
Smallwood, J., Brown, K., Baird, B., and Schooler, J. W. (2011). Samenwerking tussen het netwerk van de standaardmodus en het frontaal-pariëtale netwerk bij de productie van een interne gedachtegang. Brain Res. 1428, 60-70. doi: 10.1016 / j. brainres.2011.03.072
PubMed Abstract / CrossRef Full Text / Google Scholar
Smith, M. E., Mcevoy, L. K., and Gevins, A. (2002). De impact van matig slaapverlies op neurofysiologische signalen tijdens de working-memory taakprestaties. Slaap 25, 784-794. doi: 10.1093/sleep / 25.7.56
CrossRef Full Text / Google Scholar
Thomas, M. (2003). Neurale basis van alertheid en cognitieve prestatiestoornissen tijdens slaperigheid ii. effecten van 48 en 72 uur slaapgebrek op wakker menselijke regionale hersenactiviteit. Thalam. Relat. Syst. 2, 199–229. doi: 10.1016/S1472-9288(03)00020-7
CrossRef Full Text/Google Scholar
Van Dongen, H. (2004). De cumulatieve kosten van extra waakzaamheid: doseresponse effecten op neurologische gedragsfuncties en slaapfysiologie van chronische slaapdeprivatie en totale slaapdeprivatie. Slaap 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef Full Text / Google Scholar
Verweij, I. M., Romeijn, N., Smit, D. J., and Giovanni, P. (2014). Slaaptekort leidt tot een verlies van functionele connectiviteit in frontale hersengebieden. BMC Neurosci. 15:88. doi: 10.1186 / 1471-2202-15-88
PubMed Abstract / CrossRef Full Text / Google Scholar
Wen, X., Liu, Y., Yao, L., En Ding, M. (2013). Top-down regeling van de standaard modus activiteit in ruimtelijke visuele aandacht. J. Neurosci. 33, 6444–6453. doi: 10.1523 / JNEUROSCI.4939-12. 2013
CrossRef Full Text / Google Scholar
Wiggins, E., Malayka, M., and Kendra, G. (2018). 24-uurs slaaptekort schaadt vroege attentionele modulatie van neurale verwerking: een event-gerelateerde hersenpotentiaal studie. Neurowetenschappen. Lett. 677, 32–36. doi: 10.1016 / j.neulet.2018.04.022
PubMed Abstract / CrossRef Full Text / Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y., and Zhou, Q. (2019). Verminderde informatievervanging van werkgeheugen na slaaptekort: bewijs van een event-gerelateerde potentiële studie. Voorkant. Neurowetenschappen. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef Full Text / Google Scholar
Zhang, Q., Liao, Y., Qi, J., Zhao, Y., Zhu, T., Liu, Z., et al. (2014). Een visueel erp-onderzoek naar impulsremming na een zaleplon-geïnduceerd nap na slaaptekort. PLoS One 9: e95653. doi: 10.1371 / journal.pone.0095653
PubMed Abstract / CrossRef Full Text / Google Scholar
Leave a Reply