kirjahylly
huolimatta perifeerisestä sijainnistaan, silmän verkkokalvo eli hermoosa, on todellisuudessa osa keskushermostoa. Kehityksen aikana verkkokalvo muodostuu diencephalonin eli optisen vesikkelin ulkoreunana, joka tunkeutuu sisään ja muodostaa optisen kupin (Kuva 11.3; Katso myös Luku 22). Optisen kupin sisäseinä synnyttää verkkokalvon, kun taas Ulkoseinä synnyttää pigmenttiepiteelin. Tämä epiteeli on melaniinipitoinen rakenne, joka vähentää silmään tulevan valon takaisinsiroutumista; sillä on myös tärkeä rooli fotoreseptorien ylläpidossa, fotopigmenttien uudistamisessa ja fotoreseptorilevyjen fagosytoimisessa, koska niiden vaihtuvuus suurella nopeudella on välttämätöntä näön kannalta.

Kuva 11.3
ihmissilmän kehitys. A) verkkokalvo kehittyy hermostoputkesta lähtevänä ulkoreunana, jota kutsutaan optiseksi vesikkeliksi. (B) optinen rakkula tunkeutuu muodostaen optisen kupin. (C, D) sisäseinä optisen kupin tulee hermoston verkkokalvo, kun taas (lisää…)
sopusoinnussa sen statuksen kanssa, että verkkokalvo on keskushermoston täysivaltainen osa, se koostuu monimutkaisesta hermopiiristä, joka muuntaa fotoreseptorien luokitellun sähköisen toiminnan aktiopotentiaaleiksi, jotka kulkevat aivoihin näköhermon aksonien kautta. Vaikka verkkokalvossa on samantyyppisiä toiminnallisia elementtejä ja välittäjäaineita kuin muissa keskushermoston osissa, verkkokalvo koostuu vain muutamasta hermosoluluokasta, ja nämä ovat järjestäytyneet tavalla, jota on ollut helpompi purkaa kuin muiden aivojen piirejä. Verkkokalvossa on viittä eri hermosolutyyppiä: fotoreseptorit, kaksisuuntaiset solut, hermosolut, vaakasolut ja amakriinisolut. Näiden neuronien solurungot ja prosessit ovat pinottuina viiteen vuorottelevaan kerrokseen, solurungot sijaitsevat sisemmissä ydin -, ulompi ydin-ja hermosolukerroksissa, ja prosessit ja synaptiset kontaktit sijaitsevat sisemmissä pleksikerroksissa ja ulommissa pleksikerroksissa (Kuva 11.4). Suora kolmihermoketju-fotoreseptorisolusta kaksisuuntaiseen soluun ganglionisoluun-on tärkein informaatiovirran reitti fotoreseptoreista näköhermoon.
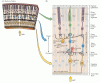
Kuva 11.4
verkkokalvon rakenne. (A) verkkokalvon osa, josta ilmenee verkkokalvon kerrosten yleinen järjestely. (B) kaavio verkkokalvon peruspiiristä. Kolmen neuronin ketju-fotoreseptori, kaksisuuntainen solu ja ganglion solu-tarjoaa suorimman (lisää…)
verkkokalvossa on kahdenlaisia valolle herkkiä elementtejä: sauvoja ja tappisoluja. Molemmilla fotoreseptorityypeillä on ulompi segmentti, joka koostuu kalvolevyistä, jotka sisältävät fotopigmentin ja sijaitsevat pigmenttiepiteelikerroksen vieressä, ja sisempi segmentti, joka sisältää solun tuman ja synnyttää synaptisia terminaaleja, jotka ottavat yhteyttä kaksisuuntaisiin tai vaakasuoriin soluihin. Valon absorptio fotoreseptorien uloimmassa segmentissä käynnistää tapahtumaketjun, joka muuttaa reseptorin kalvopotentiaalia, ja näin ollen fotoreseptorin synapsien vapauttaman välittäjäaineen määrä soluihin, joihin ne ovat yhteydessä. Fotoreseptoripäätteiden ja kaksisuuntaisten solujen (ja vaakasuorien solujen) väliset synapsit esiintyvät ulommassa pleksikerroksessa; tarkemmin sanottuna fotoreseptoreiden solurungot muodostavat uloimman ydinkerroksen, kun taas bipolaaristen solujen solurungot sijaitsevat sisemmässä ydinkerroksessa. Bipolaaristen solujen lyhyet aksonaaliset prosessit tekevät synaptisia kontakteja vuorostaan sisemmän pleksikerroksen hermosolujen dendriittisiin prosesseihin. Ganglionisolujen paljon suuremmat aksonit muodostavat näköhermon ja kuljettavat tietoa verkkokalvon stimulaatiosta muualle keskushermostoon.
verkkokalvon kahdella muulla neuronityypillä, vaakasoluilla ja amakriinisoluilla, on solurungot sisemmässä ydinkerroksessa, ja ne ovat pääasiallisesti vastuussa verkkokalvon lateraalisista vuorovaikutuksista. Nämä uloimman pleksikerroksen reseptorien, vaakasuorien solujen ja kaksisuuntaisten solujen väliset lateraaliset vuorovaikutukset ovat suurelta osin vastuussa näköjärjestelmän herkkyydestä luminanssikontrastille monenlaisilla valonvahvuuksilla. Amakriinisolujen prosessit, jotka ulottuvat sisemmän pleksikerroksen lateraalisesti, ovat postsynaptisia bipolaarisiin solupäätteisiin ja presynaptisia ganglion-solujen dendriitteihin (KS.Kuva 11.4). Horisontaalisten solujen prosessit ramiform ulompi plexiform kerros. Useita amakriinisolujen alaluokkia, jotka vaikuttavat selvästi näkökykyyn. Esimerkiksi yhdellä amakriinisoluluokalla on tärkeä rooli bipolaaristen solujen valovasteen muuttamisessa joidenkin hermosolutyyppien lyhytkestoisiksi ohimeneviksi vasteiksi. Toinen tyyppi toimii pakollisena vaiheena reitissä, joka välittää tietoa rod-fotoreseptoreista verkkokalvon ganglionisoluihin. Amakriinisolun alatyyppien moninaisuus kuvaa yleisempää sääntöä, että vaikka verkkokalvon perussolutyyppejä on vain viisi, tietyn solutyypin sisällä voi olla huomattavaa monimuotoisuutta. Tämä moninaisuus on perusta poluille, jotka välittävät erilaisia tietoja keskeisiin kohteisiin rinnakkain.
ensi silmäyksellä verkkokalvokerrosten tilajärjestely näyttää vastavaikuttavalta, sillä valonsäteiden täytyy kulkea verkkokalvon (ja verkkokalvon verisuoniston) ei-valoherkkien elementtien läpi!) ennen kuin ne saavuttavat fotoreseptorien uloimmat segmentit, joihin fotonit absorboituvat (KS.Kuva 11.4). Syynä tähän verkkokalvon organisaation omituiseen ominaisuuteen on erityinen suhde, joka vallitsee fotoreseptorien uloimpien segmenttien ja pigmenttiepiteelin välillä. Ulommissa segmenteissä on kalvolevyjä, joissa on valoherkkä valopigmentti ja muita transduktioprosessiin osallistuvia proteiineja. Nämä välilevyt muodostuvat lähelle fotoreseptorin sisäsegmenttiä ja siirtyvät kohti ulomman segmentin kärkeä, jossa ne irtoavat. Pigmenttiepiteelillä on keskeinen rooli kuluneiden reseptorilevyjen poistamisessa; tämä ei ole mikään pieni tehtävä, sillä kaikki ulompien segmenttien levyt vaihdetaan 12 päivän välein. Lisäksi pigmenttiepiteelissä on biokemiallinen koneisto, jota tarvitaan fotopigmenttimolekyylien uudistamiseen niiden altistuttua valolle. Se on oletettavasti vaatimukset photoreceptor levyn elinkaaren ja photopigment kierrätys, jotka selittävät, miksi sauvat ja käpyjä löytyy uloimmassa eikä sisin kerros verkkokalvon. Pigmenttiepiteelin ja verkkokalvon fotoreseptorien välisten normaalien suhteiden häiriöillä, kuten retinitis pigmentosa-bakteerin aiheuttamilla häiriöillä, on vakavia seurauksia näkökykyyn (laatikko B).
Box B
Retinitis Pigmentosa.
Leave a Reply