Frontiers in Neuroscience
Úvod
S rozvojem společnosti a změnami v rytmus práce, zvyšující se počet lidí, kteří trpí nedostatkem spánku. Nedostatek spánku poškozuje nejen fyzické a duševní zdraví jednotlivce, ale také vážně ovlivňuje pracovní výkon, způsobuje pracovní chyby a dokonce i nehody. Pochopení mechanismu deprivace spánku, který ovlivňuje kognitivní funkce, má proto velký význam pro účinnou prevenci účinků deprivace spánku.
předchozí studie ukázaly, že deprivace spánku může způsobit řadu změn v náladě jednotlivce, kognitivních schopnostech, pracovním výkonu a imunitní funkci (Choo et al., 2005). Nedostatek spánku narušuje cirkulaci těla a ovlivňuje kognitivní a emoční schopnosti jednotlivců(Raymond, 1988). Několik studií ukázalo, že deprivace spánku zhoršuje inhibici reakce (Harrison a Horne, 1998; Muzur et al., 2002; Jennings et al., 2003). Například po 36 hodinách deprivace spánku se snížila schopnost jedince potlačit negativní podněty (Chuah et al ., 2006). Neuroimagingové studie naznačují, že deprivace spánku snižuje nízkou úroveň schopnosti vizuálního zpracování jedince (Anderson a Platten, 2011; Ning et al ., 2014). Kromě toho deprivace spánku narušuje hipokampus a může ovlivnit paměť ničením synaptické plasticity (Cote et al ., 2014). Thomas (2003) naznačil, že nedostatek spánku snižuje průtok krve mozkem a rychlost metabolismu v thalamu, prefrontální kůře a parietální kůře (Géraldine et al ., 2005). Jarraya a jeho kolegové zjistili, že částečná deprivace spánku významně ovlivnila neuropsychologické funkce, jako je verbální okamžitá paměť, pozornost a bdělost (Thomas, 2003). Některé studie navíc odhalily, že kumulativní účinky částečné deprivace spánku by mohly vážně narušit kognitivní funkce a chování (Van Dongen, 2004; Scott et al., 2006; Jarraya a kol., 2013).
pracovní paměť je systém, který slouží k ukládání a zpracování informací a který je kognitivní funkcí s omezenou kapacitou (Bartel et al ., 2004). Kromě toho mohou být informace uložené v systému pracovní paměti změněny z krátkodobé paměti na dlouhodobou paměť pomocí retellingu a dalších metod paměti. Pracovní paměť je přechod mezi krátkodobými a dlouhodobými paměťovými systémy, který je velmi klíčový při zpracování lidských zpráv (Miyake and Shah, 1999). Poskytuje dočasný úložný prostor a zdroje potřebné ke zpracování informací, jako je porozumění hlasu, uvažování a učení. Ukázalo se, že deprivace spánku nejprve ovlivňuje pracovní paměť.
Předchozí studie použili n-back paradigmatu pracovní paměti u účastníků, kteří podstoupili spánku a zjistil, že nedostatek spánku způsobuje pokles metabolické aktivity v mozku je regionální síť, která je převážně provedeno zpracování informací a reakce inhibice (Baddeley, 2000; Zhang et al., 2019). Zhoršená pracovní paměť po deprivaci spánku souvisí s aktivací výchozí sítě v úkolech (Chee a Chuah, 2008), což může souviset s důležitou rolí thalamu v kortikální bdělosti. Například, nedostatek spánku zvyšuje spojení mezi hipokampu, thalamu, a výchozí síť, která byla často doprovázena vyšší subjektivní ospalost a horší výkon pracovní paměti (Lei et al., 2015; Li a kol., 2016). Studie o spánkové deprivaci?ed, že zvýšená latence a snížená amplituda složky P3 byly spojeny s prodlouženou střízlivostí (Morris et al ., 1992; Jones a Harrison, 2001; Panjwani a kol., 2010). Snížení P3 vlna může odrážet pokles účastníků pozornosti a snížení rozlišování cílových podnětů (Koslowsky a Babkoff, 1992).
několik studií však poskytlo elektrofyziologické důkazy o zhoršené pracovní paměti po deprivaci spánku. Úloha n-back je považována za běžnou metodu hodnocení pracovní paměti (Owen et al ., 2005; Jaeggi a kol., 2010). Zhang et al. navrhl dvě zadní výslovnost pracovní paměť úkolem prozkoumat snížil zpráva alternativní pracovní paměti během spánku, ale jen málo studií používá různé druhy pracovní paměti úkoly v jediném experimentu. V této studii jsme navrhli různé typy pracovní paměť úkolů (výslovnost, pracovní paměť, prostorovou pracovní paměť, a objekt pracovní paměť), aby prozkoumala zhoršení kognitivní funkce tím, TSD a zaznamenal účastník EEG dat na 2 časové body (výchozí a 36 h-TSD). Všechny úkoly přijaly paradigma 2-back. Tato studie hodnotila změny ve vlně N2-P3 související s pracovní pamětí během TSD a analyzovala časové charakteristiky účinků deprivace spánku na pracovní paměť. Naše zjištění poskytují experimentální důkazy o účincích deprivace spánku na kognitivní funkce.
materiály a metody
účastníci
této studie se zúčastnilo šestnáct mladých, zdravých, praváků. Rekrutovali jsme účastníky reklamou na kampusu. Všichni účastníci měli dobré spánkové návyky (PSQI< 5). Všichni účastníci byli ve věku mezi 21 a 28 let s průměrným věkem 23 let, a žádný z účastníků neměl žádné psychické nebo fyzické nemoci. Všichni účastníci měli normální vidění nebo korigované vidění nad 1,0 a skóre inteligence > 110 V Ravenově testu. Před experimentem experimentátor vysvětlil postup a upozornil účastníky, aby se ujistili, že jsou obeznámeni s metodou a postupem. V 2 týdny před zahájením experimentu účastníci spali pravidelně pro 7-9 h denně, bez kouření, pití kávy, pití alkoholu, nebo konzumovat žádné léky 2 dny před experimentem. Před experimentem poskytli všichni účastníci písemný informovaný souhlas. Experimentální schéma bylo schváleno etickou komisí čtvrté Vojenské lékařské univerzity a univerzity Beihang.
experimentální návrh
všem účastníkům byly představeny tři typy úloh pracovní paměti. Byly to dva-back výslovnost pracovní paměti úkol (viz Obrázek 1), dva-back prostorové pracovní paměti úkol (viz Obrázek 2), a dva-back objekt pracovní paměti úkol (viz obrázek 3). Stimulačními materiály úkolů bylo 15 písmen necitlivých anglických písmen, která kromě těch s podobnými písmeny, jako jsou L/ l, M / m; malé černé čtverečky; a 12 geometrických obrazců. Všechny materiály byly zobrazeny v černé barvě na bílém pozadí s přibližným zorným úhlem 1,5 ° × 1,5° (šířka: 2,0 cm, výška: 2,0 cm). 122 stezky byly obsaženy v každý úkol a každou stopu, cílový stimul byl prezentován po dobu 400 ms dvě stezky po předložení objektivních stimulů, s 1600 ms stimul nástupem nejsou synchronizovaná čas (SOA), která byla označena bílý „+.“Účastníci byli požádáni, aby klikněte na levé tlačítko myši, když je cíl a cíl podněty byly stejné („matching“), zatímco klepnutím na pravé tlačítko myši když nebyly („nesoulad“). Podmínka shody nebo ne byla prezentována v pseudonáhodném pořadí s poměrem 1: 1.
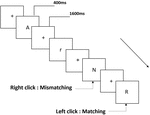
Obrázek 1. Schematické schéma úlohy pracovní paměti výslovnosti.
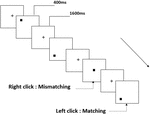
Obrázek 2. Schéma úlohy prostorové pracovní paměti.
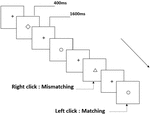
Obrázek 3. Schéma úlohy pracovní paměti objektu.
experimentální postupy
před experimentem byli účastníci poučeni o experimentálním úkolu. Byli informováni, aby procvičovali tři typy úloh pracovní paměti, dokud nebylo dosaženo míry přesnosti 90%. Účastníci navštívili laboratoř den před experimentem a tu noc spali v laboratoři. Oba partnerští účastníci provedli experimenty současně. Tři typy úloh pracovní paměti byly provedeny v 7:30 do 8:30 následujícího rána se současným záznamem elektroencefalogramu (EEG) (základní linie). Druhý záznam EEG (36 h-TSD) byl proveden po období 36 hodin, během kterého účastníci nemohli spát. Během celé doby experimentu byly zakázány centrální inhibice a stimulační léky. Účastníci byli spolu, pozorovat a připomněl ošetřovatelského personálu s cílem udržet je vzhůru celé TSD zasedání.
EEG nahrávky
kontinuální skalp EEG byl zaznamenán pomocí uzávěrů elektrod umístěných na 64 místech pomocí systému 10-20 se zesilovačem SynAmps2. Pro referenci byly použity bilaterální mastoidy (A1 a A2) a čelo bylo použito jako zem. EEG byly zaznamenány při 1000 Hz a impedance všech kanálů byla udržována pod 5 kΩ. Další čtyři elektrody byly umístěny nad a pod pravé a levé oči pro záznam bipolární vertikální a horizontální electrooculogram.
Analýza dat behaviorálních experimentů
kvůli technickým chybám byly dva případy odstraněny, zatímco dalších 14 případů bylo zahrnuto do následující statistické analýzy. Údaje o chování zahrnovaly střední reakční dobu, správnou rychlost a správný počet za jednotku času. Behaviorální údaje ve výchozím stavu a 36 stavy h-TSD byly zaznamenány pro analýzu. Analýzy byly prováděny společností IBM SPSS (V22.2), kde byla zahájena post-hoc analýza opakovaných měření rozptylu (ANOVA) metodou se skleníkem-Geisser Bonferroni. Statistické výsledky byly prezentovány jako průměr a směrodatná odchylka (SD).
Analýza dat EEG
Scan 4.3 program byl použit k analýze dat EEG, kde byly EEG artefakty pohybu očí korigovány metodou redukce očních artefaktů. Epochy v rozmezí od -100 do 800 ms kontinuálních dat EEG byly extrahovány a filtrovány pásmovým filtrem od 0,5 do 30 Hz s frekvenčním sklonem 24 dB / oct. V klinických studiích, v nichž napětí překročila ± 100 µV byly zamítnuty a základní byl opraven na mysli amplituda 100 ms. EEG komponenty byly zprůměrovány a vypočteny pouze s opravenou odpovědí. Byly identifikovány a kvantifikovány komponenty ERP P2 (100-250 ms), N2 (150-350 ms) a P3 (250-450 ms) stimulačních studií. Grand-průměrná hodnota maximální amplitudy a latence N2 a P3 komponenty byly vypočteny odděleně na F3 Fz F4, C3, Cz, C4, P3, Pz, P4 a P2 součástí byla vypočtena na F3 Fz F4, C3, Cz, a C4 (Špaletová et al., 2006; Verweij et al., 2014).
opakovaná opatření ANOVA byla použita pro všechny výsledky ERP. Hlavní efekty a interakce mezi režimy spánku (základní a 36 h-TSD), úkoly (výslovnost, pracovní paměť, prostorovou pracovní paměť, a objekt pracovní paměť), regiony (frontální, centrální, parietální; P2 součástí byla analyzována pouze na čelní a centrální regiony), a stránky (vlevo, uprostřed, vpravo) byly statisticky analyzovány zaměstnávání opakovaná měření ANOVA, která zahrnovala Greenhouse-Geisser korekce pro non-kulovitého tvaru a Bonferroniho post-hoc testy.
Výsledky
Behaviorální Výkon
výsledky behaviorálních experimentů jsou uvedeny v Tabulce 1. Průměrná reakční doba byla delší ve stavu 36 h-TSD než na začátku s trendem ke zvýšení, ale bez významných rozdílů. ANOVA odhalila, že správná rychlost úkolu byla významně odlišná mezi základní linií a 36 h-TSD . Správný počet za jednotku času vykazoval významný hlavní účinek času během 36 h-TSD .

Tabulka 1. Údaje o výkonu (průměr ± SD) na úkolu 2-back na začátku a po 36 h-TSD.
amplituda
ve srovnání s výchozí hodnotou bylo pozorováno významné snížení amplitudy P3 a významné zvýšení amplitudy P2 po TSD. Ačkoli amplituda N2 klesla po 36 hodinách TSD, rozdíl nedosáhl statistické významnosti (Tabulka 2).

Tabulka 2. Velká průměrná špičková amplituda složek P2, N2 a P3 ve správném stavu odezvy na více elektrodových místech na začátku a po 36 h-TSD.
byly zjištěny významné hlavní účinky oblastí a míst na amplitudu P2 za podmínky TSD. Během TSD se v čelní oblasti objevila maximální amplituda P2 (obrázek 4). Kromě toho, rozdíly v P2 amplitud v různých regionech (čelní vs. centrální) byly výraznější v bilaterální elektrody (vlevo: P = 0,001; pravý: P = 0.000), než ve střední elektrody (Obrázek 4). Významný hlavní efekt pro region a místo v N2 bylo zjištěno, že N2 amplituda byla více negativní v čelní, než v centrální oblasti (P = 0.008, Obrázek 4) a menší na pravé straně než na levé straně (P = 0.011, Postavy, 5A2,B2). Byl pozorován hlavní účinek místa na amplitudu P3 . Amplituda P3 byla ve středu pozitivnější než na levé straně (P = 0,009, obrázek 4). Pro amplitudu P3 byl pozorován významný interakční účinek mezi časem a oblastí . Během TSD bylo snížení amplitudy P3 významnější ve frontálních a centrálních oblastech než v parietální oblasti (P = 0,005; P = 0,003) (obrázek 5C3). Žádné další hlavní účinky nebo interakční účinky nedosáhly statistické významnosti.
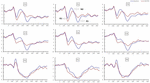
obrázek 4. Amplituda ERP na začátku a 36 h-TSD pro správnou podmínku odezvy pro úlohu pracovní paměti. Kanály jsou seřazeny zleva doprava a shora dolů takto: F3, Fz, F4, C3, Cz, C4, P3, Pz a P4. Ve srovnání s výchozí hodnotou byly latence složek N2-P3 prodlouženy a amplitudy N2-P3 byly sníženy po 36 h-TSD.
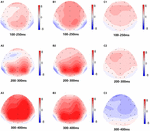
Obrázek 5. Topografická mapa správné odezvy v úloze pracovní paměti v různých podmínkách spánku (A1-C3). (A1) P2, 100-250 ms, na začátku. (A2) N2, 200-300 ms, na začátku. (A3) P3, 300-400 ms, na začátku. (B1) P2, 100-250 ms, při 36 h-TSD. (B2) N2, 200-300 ms, pro 36 h-TSD. (B3) P3, 300-400 ms, pro 36 h-TSD. (C1) P2, 100-250 ms, 36 h-TSD se odečtenou základní hodnotou. (C2) N2, 200-300 ms, 36 h-TSD se odečtením základní linie. (C3) P3, 300-400 ms, 36 h-TSD se odečtenou základní hodnotou.
Latence
latence N2 a P2 byly výrazně delší po TSD. Ačkoli latence P3 byla prodloužena po 36 hodinách TSD, rozdíl nedosáhl statistické významnosti (Tabulka 3).

Tabulka 3. Grand-průměrná maximální latence složek P2, N2 a P3 ve správném stavu odezvy na více elektrodových místech na začátku a po 36 h-TSD.
významný hlavní účinek regionu na N2 a P3 bylo zjištěno, že latence N2-P3 komponenty byla kratší v temenní oblasti, než v čelní oblasti (P = 0.002; P = 0.000) a středočeský kraj (P = 0.000; P = 0.000) (Obrázek 4). Latence vlny P3 byla výrazně delší na levé straně než na pravé straně (Obrázek 4).
žádné další hlavní účinky nebo interakční účinky nedosáhly statistické významnosti.
amplitudy a latence N2, P2 a P3, které byly vyvolány na devíti elektrodových místech, jsou uvedeny na obrázku 4. Topografická mapa správná odpověď v pracovní paměť úkolem v různé spát podmínky (základní, 36 h-TSD a rozdíl mezi dvou podmínek) je prezentován na Obrázku 5.
Diskuse
V této studii, jsme informovali vlivům 36 h spánku na pracovní paměť, kombinuje behaviorální data ve dvou spát státy (základní a 36 h-TSD) s soudobé záznamy EEG. Z analýzy výsledků vyplynulo, že změny v chování dat v souladu s poruchou pracovní paměti po 36 h TSD: zvýšení průměrné reakční doba kognitivní úkoly a snížení přesnosti.
deprivace spánku narušila individuální kontrolu nad zdroji pozornosti. I když jedinci se snažil udržet bdělost a výkon práce, včetně reakční čas a správnou rychlost, během spánku, informace zpracovatelské kapacity jejich pracovní paměti byl ještě ovlivněn, protože pokles rychlosti zpracování informací (Špaletová et al., 2006; Wiggins a kol., 2018). V této studii, N2 a P3 vlny týkající se pracovní paměť byla měřena na zvýšení latence a snížení amplitudy po spánku ve srovnání s výchozí hodnoty. Studie prokázaly, že nedostatek spánku vede k neustálému poklesu pozornosti, a jev, snížení P3 amplituda ukazuje, že jedinci,‘ top-down control poznání se postupně hroutí. Deprivace spánku má nepříznivější účinek na kognitivní funkce, zejména ty, které závisí na duševních nebo kognitivních funkcích (Kusztor et al ., 2019).
komponenta P3 odráží nasazení prostředků pozornosti a latence P3 je široce považována za časové okno pro kategorizaci a hodnocení stimulů. Snížení amplitudy vlny P3 také potvrdilo, že rozhodování v odpovídající odpovědi po TSD bylo do určité míry poškozeno (Gosselin et al., 2005). Studie naznačují, že deprivace spánku může ovlivnit fázi zpracování informací pracovní paměti. V této studii ukazatele výkonnosti také podpořily závěr, že doba odezvy na cílový stimul byla zvýšena a že latence vlny P3 byla prodloužena (Cote et al., 2008). To bylo spekuloval, že vliv spánku na P3 součástí může také probíhat, protože neschopnost reagovat na informace měnit, což je v souladu s předchozími závěry, že P3 komponenty se vztahují k aktualizaci pracovní paměti obsah (Donchin a Fabiani, 1991).
předchozí studie považovaly složku N2 za elektrofyziologický index odrážející schopnost jedince potlačit odpověď (Kreusch et al., 2014). Po deprivaci spánku prodloužená latence složky NoGo-N2 naznačuje, že schopnost jedince potlačit odpověď je narušena (Jin et al ., 2015). Snížená amplituda a prodloužená latence složky N2 související s pracovní pamětí výslovnosti po deprivaci spánku odhaluje, že deprivace spánku narušuje zpracování informací o pracovní paměti výslovnosti (Zhang et al ., 2019). Obecně se předpokládá, že složka N2 odráží selektivní pozornost mozku a zpracování emočních podnětů nebo signálů (Schacht et al., 2008) a je endogenní složkou související s duševním stavem, pozorností a stupněm pozornosti jednotlivce. V této studii jsme zjistili, že latence složky N2 byla významně prodloužena, ale amplituda vykazovala pouze klesající trend. Podle předchozích studií prodloužená latence N2 odrážela prodloužení doby odezvy po omezení spánku (Zhang et al ., 2014). Zjištění, že amplituda N2 nebyla významně změněna, však mohlo být způsobeno mozkovými kompenzačními odpověďmi (Drummond a Brown, 2001). V případě omezených kognitivních zdrojů existoval kompenzační mechanismus pro obnovení zhoršené kognitivní funkce (Jin et al ., 2015).
podle topografie pokožky hlavy jsou změny složek N2-P3 související s deprivací spánku zřetelnější v čelní oblasti. Frontoparietální kontrola (FPC) hraje důležitou roli v kognitivní kontrole. Studie ukázaly, že FPC může obejít kognitivní kontrolu shora dolů, což umožňuje jednotlivcům soustředit se na informace týkající se cíle a zároveň potlačovat informace, které nesouvisejí s cílem (Smallwood et al ., 2011; Wen et al., 2013). FPC je důležité pro uchování informací a zpracování informací v pracovní paměti, a stupeň aktivace FPC po spánku byla snížená ve srovnání s normální spánek skupina (Ma et al., 2014). Ačkoli výsledky EEG neodrážely změny v konkrétních oblastech mozku podrobně, intuitivně odrážely účinek TSD na uchovávání a zpracování informací o pracovní paměti.
i když přesný kognitivní proces, který P2 součást základem je stále široce diskutován, jako široká definice, P2 složka odráží proces, pozornosti a vizuální zpracování a je obecně považováno za vztahující se k selektivní pozornosti a pracovní paměti, což odráží rané rozsudek percepční proces (Saito et al., 2001). V této studii jsme zjistili významné zvýšení amplitudy vlny P2 po deprivaci spánku. Studie uvádějí, že P2 vlny, které by mohly být součástí systému raného kognitivního párování pro zpracování zpráv a mohou porovnávat smyslové vstupy s uloženou pamětí (Freunberger et al ., 2007) jsou citliví na změny v pozornosti mise a požadavcích na pracovní paměť (Smith et al., 2002). Funkční kompenzace je jednou z jedinečných funkcí lidského mozku a důležitým faktorem pro udržení kognitivní funkce. Nadměrné aktivace dorzolaterální prefrontální kortex (DLPFC) po spánku naznačuje, že, jako mozek zdrojů snížit, DLPFC se zdá, že má kompenzační funkci (Drummond et al., 2004; Choo et al., 2005). Proto spekulujeme, že významné zvýšení amplitudy P2 pozorované v této studii může být způsobeno funkční kompenzací, ve které se zdá, že jednotlivci udržují normální kognitivní funkce po deprivaci spánku. I když velké množství studií používá ERP technologie, aby prozkoumala vliv nedostatku spánku na kognitivní funkce, brzy komponent, jako jsou N1 a P2 nebyly systematicky studovány, a výsledky jsou nekonzistentní (Evans a Federmeier, 2007; Wiggins et al., 2018; Zhang et al., 2019). Existuje jen málo výzkumů, které zkoumají změnu složky P2 během deprivace spánku (Mograss et al ., 2009). Proto je třeba účinky deprivace spánku na rané složky ERP, jako je P2, ještě dále studovat a prozkoumat.
v tomto experimentu jsme použili model 2-back k návrhu úloh výslovnosti, prostorové a objektové pracovní paměti a zkoumali poškození pracovní paměti po 36 hodinách TSD. Ve srovnání s předchozí studií, která se zaměřila pouze na účinky spánku na určitý typ informací, jako jsou výslovnost, pracovní paměť, nebo specifické kognitivní funkce, jako reakci inhibice, zvažovali jsme obsah pracovní paměti modelu a komplexně analyzován vliv spánku na pracovní paměť.
studie má však určitá omezení. Nejprve jsme použili pouze úlohu 2-back a nedokázali jsme porovnat výkon účastníků v úlohách pracovní paměti s různými obtížemi. Proto existují omezení při vysvětlování a odvozování změn v pracovní zátěži. Za druhé, ve studii byly použity pouze dobrovolníci mužského pohlaví a závěry je třeba posoudit při jejich rozšíření na dobrovolnice. Vzhledem k omezenému počtu účastníků jsme zjistili pouze to, že amplituda vlny N2 měla sestupný trend a že latence vlny P3 měla prodloužený trend. Stabilní výsledky lze získat po zvýšení počtu účastníků. Zatřetí, kombinace našeho postupu s fMRI pro pracovní paměť může usnadnit další interpretaci výsledků. Předchozí studie ukázaly, že cirkadiánní biorytmy ovlivňují behaviorální výkon a existují individuální rozdíly (Montplaisir, 1981; Lavie, 2001). V tomto experimentu jsme nezaznamenali data EEG ve stejném časovém bodě, takže vliv cirkadiánních biorytmů na výsledky testů nelze zcela vyloučit.
tento výzkum ukázal, že schopnost pracovní paměti byla po TSD narušena a že toto poškození nebylo spojeno s stimulačním obsahem pracovní paměti. Nedostatek spánku snižuje kvalita informací uložených v paměti, které mohou nastat s degenerativní proces pozornosti (Ratcliff a Van Dongen, 2018). Tato studie poskytuje elektrofyziologické důkazy pro pochopení mechanismu pod zhoršenou pracovní pamětí po deprivaci spánku. Je třeba věnovat pozornost nepříznivým účinkům poruchy pracovní paměti způsobené deprivací spánku a prozkoumat účinné zásahy do takového poškození.
Prohlášení o dostupnosti dat
datové sady vytvořené pro tuto studii jsou k dispozici na vyžádání příslušnému autorovi.
etické prohlášení
studie zahrnující lidské účastníky byly přezkoumány a schváleny čtvrtou Vojenskou lékařskou univerzitou Beihang University. Pacienti / účastníci poskytli písemný informovaný souhlas s účastí v této studii.
Autor příspěvky
YS navrhl experimenty. ZP produkoval výsledky a napsal rukopis. CD a LZ analyzovaly a interpretovaly data. JT a YS provedli experimenty, seznámili se s daty a garanty této studie. YB, LZ a JT přispěly k účasti na sběru dat a revizi literatury. Všichni uvedení autoři přečetli a schválili konečný rukopis.
financování
tento výzkum byl podpořen Národní vědeckou nadací technologického plánu Zimních olympijských her v Číně v rámci grantu nos. 2019YFF0301600 a HJ20191A020135.
střet zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být vykládány jako potenciální střet zájmů.
Anderson, C., and Platten, C. R. (2011). Deprivace spánku snižuje inhibici a zvyšuje impulzivitu k negativním podnětům. Chovej se. Brain Res.217:466. doi: 10.1016 / j. bbr.2010.09.020
PubMed Abstract / CrossRef Full Text / Google Scholar
Baddeley, a. (2000). Epizodická vyrovnávací paměť: nová součást pracovní paměti? Trendy Cogn. Věda. 4:417. doi: 10.1016/s1364-6613(00)01538-2
CrossRef Plný Text | Google Scholar
Bartel, P., Offermeier, W., Smith, F., a Becker, P. (2004). Pozornost a pracovní paměť u rezidentních anesteziologů po noční službě: skupinové a individuální účinky. Okupace. Environ. Med. 61, 167–170. doi: 10.1136 / oem.2002.006197
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Křídla, M. D., Broussard, J. L., Mullington, J. M., a Stiskněte tlačítko D. Z. (2006). Příspěvek spánku ke zlepšení rychlosti skenování pracovní paměti: studie dlouhodobého omezení spánku. Biol. Psychol. 72:212. doi: 10.1016 / j. biopsycho.2005.11.002
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Chee, M. W. L., a Chuah, L. Y. M. (2008). Funkční neuroimaging vhled do toho, jak spánek a deprivace spánku ovlivňují paměť a poznání. Curre. Opine. Neurol. 21, 417–423. doi: 10.1097 / WCO.0b013e3283052cf7
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S., a Chee, M. W. L. (2005). Disociace kortikálních oblastí modulovaných jak zatížením pracovní paměti, tak deprivací spánku a samotnou deprivací spánku. Neuroimage 25, 579-587. doi: 10.1016 / j. neuroimage.2004.11.029
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Chuah, Y. M. L., Venkatraman, V., Dinges, D. F., a Chee, M. W. L. (2006). Neurální základ interindividuální variability v inhibiční účinnosti po deprivaci spánku. J. 26, 7156–7162. doi: 10.1523/jneurosci.0906-06.2006
CrossRef Plný Text | Google Scholar
Cote, K. a., Milner, C. E., Osip, S. L., Baker, M. L., a Cuthbert, B. P. (2008). Fyziologické vzrušení a pozornost během týdne nepřetržitého omezení spánku. Fyziol. Chovej se. 95, 353–364. doi: 10.1016 / j. physbeh.2008.06.016
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Cote, K. a., Mondloch, C. J., Sergeeva, v., Taylor, M., and Semplonius, T. (2014). Vliv celkové spánkové deprivace na behaviorální nervové zpracování emocionálně výrazných tváří. Expo. Brain Res.232, 1429-1442. doi: 10.1007/s00221-013-3780-1
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Donchin, E., a Fabiani, M. (1991). Využití mozkových potenciálů souvisejících s událostmi ve studiu paměti: je p300 měřítkem rozlišovací způsobilosti událostí? Psychol. Facul. Veřejnost. 7, 58–77. doi: 10.1111 / sltb.12055
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Drummond, S. P., Brown, g. G., Salamat, J. S., a Gillin, J. C. (2004). Zvyšující se obtížnost úkolu usnadňuje mozkovou kompenzační reakci na celkovou deprivaci spánku. Spánek 27, 445-451. doi: 10.1093/spánek/27.3.445
CrossRef Plný Text | Google Scholar
Drummond, S. P. a., Brown, g. G. (2001). Účinky celkové deprivace spánku na mozkové reakce na kognitivní výkon. Neuropsychofarmakologie 25, S68-S73. doi: 10.1016/S0893-133X(01)00325-6
CrossRef Plný Text | Google Scholar
Evans, K. M., a Federmeier, K. D. (2007). Paměti, že\“pravdu a paměť, která\“zbylo: event-related potenciálů odhalit hemisferální asymetrie v kódování a uchovávání verbální informace. Neuropsychologie 45, 1777-1790. doi: 10.1016 / j. neuropsychologie.2006.12.014
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Freunberger, R., Klimesch, W., Doppelmayr, M., a Höller, Y. (2007). Vizuální složka p2 souvisí s fázovým blokováním theta. Neurovědci. Lette. 426, 181–186. doi: 10.1016 / j. neulet.2007.08.062
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Géraldine, R., Desgranges, B., Foret, J., a Eustache, F. (2005). Vztahy mezi paměťovými systémy a fázemi spánku. J. Sleep Res.14, 123-140. doi: 10.1111 / j. 1365-2869. 2005. 00450.x
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Gosselin, A., Koninck, J. D., Campbell, K. B. (2005). Celková deprivace spánku a zpracování novinek: důsledky pro fungování čelního laloku. Cline. Neurofyziol. 116, 211–222. doi: 10.1016 / j. clinph.2004.07.033
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Harrison, Y., Hornová, J. (1998). Ztráta spánku narušuje krátké a nové jazykové úkoly s prefrontálním zaměřením. J. Sleep Res.7, 95-100. doi: 10.1046 / j. 1365-2869. 1998. 00104.x
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Jaeggi, S. M., Buschkuehl, M., Perrig, W. J., a Meier, B. (2010). Souběžná platnost úlohy N-back jako opatření pracovní paměti. Paměť 18, 394-412. doi: 10.1080/09658211003702171
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Jarraya, S., Jarraya, M., Chtourou, H., Souissi, N., a Chamari, K. (2013). Dopad částečné deprivace spánku na denní variace kognitivního výkonu u vyškolených subjektů. Proc. SOC. Chovej se. Věda. 82, 392–396. doi: 10.1016 / j. sbspro.2013.06.281
CrossRef Plný Text | Google Scholar
Jennings, J. R., Monk, T. H., a Molen, M. W. V. D. (2003). Deprivace spánku ovlivňuje některé, ale ne všechny procesy dohledu. Psychol. Věda. 14, 473–479. doi: 10.1111/1467-9280.02456
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Jin, X, Ye, E., Qi, J., Wang, L., Lei, Y. (2015). Zotavení spánek zvrátí zhoršenou inhibici odezvy v důsledku omezení spánku: důkazy ze studie potenciálů souvisejících s vizuální událostí. PLoS One 10:e0142361. doi: 10.1371 / deník.pone.0142361
PubMed Abstract / CrossRef Full Text / Google Scholar
Jones, K., and Harrison, y. (2001). Funkce čelního laloku, ztráta spánku a fragmentovaný spánek. Spánková Medička. Rev. 5: 475. doi: 10.1053/smrv.2001.0203
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Koslowsky, M., a Babkoff, H. (1992). Metaanalýza vztahu mezi celkovou deprivací spánku a výkonem. Chronobiol. Int. 9, 132–136. doi: 10.3109/07420529209064524
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Kreusch, F., Quertemont, E., Vilenne, A., a Hansenne, M. (2014). Zneužívání alkoholu a komponenty erp v úkolech go/no-go pomocí podnětů souvisejících s alkoholem: dopad vyhýbání se alkoholu. Int. J. Psychofyziol. 94, 92–99. doi: 10.1016 / j. ijpsycho.2014.08.001
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Kusztor, A., Raud, L., Juel, B. E., Nilsen, A. S., Storm, J. F., a Huster, R. J. (2019). Deprivace spánku odlišně ovlivňuje subkomponenty kognitivní kontroly. Spánek 42: zsz016. doi: 10.1093/spánek/zsz016
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Lavie, P. (2001). Spánek-probuzení jako biologický rytmus. Annu. Reverend Psychol. 52, 277–303. doi: 10.1146 / annurev.psycha.52.1.277
CrossRef Full Text / Google Scholar
Lei, y., Shao, y., Wang, L., Zhai, T., and Zou, F. (2015). Rozsáhlé propojení mozkové sítě předpovídá celkové účinky deprivace spánku na kognitivní kapacitu. PLoS One 10: e0133959. doi: 10.1371 / deník.pone.0133959
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Li, C., Huang, D., Qi, J., Chang, H., a Meng, Q (2016). Krátkodobé deficity paměti korelují se změnami funkční konektivity hippocampal-thalamic po akutním omezení spánku. Zobrazování Mozku Se Chová. 11, 954–963. doi: 10.1007/s11682-016-9570-1
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Ma, N., Dinges, D. F., Basner, M., a Rao, H. (2014). Jak akutní celková ztráta spánku ovlivňuje ošetřující mozek: metaanalýza neuroimagingových studií. Prospal 38, 233. doi: 10.5665 / spánek.4404
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Miyake, A., Šach, P. (1999). Modely pracovní paměti. Mechanismy aktivní údržby a výkonné kontroly. Cambridge: Cambridge University Press.
Google Scholar
Mograss, M. a., Guillem, F., Brazzini-Poisson, V., a Godbout, R. (2009). Účinky celkové deprivace spánku na procesy paměti rozpoznávání: studie potenciálu souvisejícího s událostmi. Neurobiol. Naučit. Paměť. 91, 343–352. doi: 10.1016 / j. nlm.2009.01.008
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
mon plaisir, J. (1981). Deprese a biologický rytmus: terapeutické účinky deprivace spánku. L ‚ Union Médicale du Canada 110, 272-276. doi: 10.1620 / tj.133.481
PubMed Abstract / CrossRef Full Text / Google Scholar
Morris, A. M., So, y., Lee, K. a., Lash, a. A., a Becker, C. E. (1992). Potenciál související s událostmi p300. účinky deprivace spánku. J. Med. 34, 1143–1152.
Google Scholar
Muzur, a., Pace-Schott, E. F., and Hobson, J. a. (2002). Prefrontální kůra ve spánku. Trendy Cogn. Věda. 6, 475–481. doi: 10.1016/S1364-6613(02)01992-7
CrossRef Plný Text | Google Scholar
Ning, M. Dinges, D. F., Matyáš, B., a Hengyi, R. (2014). Jak akutní celková ztráta spánku ovlivňuje ošetřující mozek: metaanalýza neuroimagingových studií. Spánek 38, 233-240. doi: 10.5665 / spánek.4404
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Owen, a. M. Mcmillan, K. M., Zeman, A. R., a Bullmore, E. (2005). Paradigma pracovní paměti N-back: metaanalýza normativních funkčních neuroimagingových studií. Hučení. Brain Mapp. 25, 46–59. doi: 10.1002 / hbm.20131
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Panjwani, U., Ray, K., Chatterjee, A., Bhaumik, a. S., a Kumar, S. (2010). Elektrofyziologické koreláty poznávání se zlepšují s zdřímnutím během deprivace spánku. Euro. J.Appl. Fyziol. 108, 549–556. doi: 10.1007/s00421-009-1222-3
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Ratcliff, R., a Van Dongen, H. (2018). Účinky deprivace spánku na položku a asociativní rozpoznávací paměť. J.Exp. Psychol. 44, 193–208. doi: 10.1037/xlm0000452
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Raymond, C. a. (1988). Přesouvání práce, spánkové cykly jsou na cestě k tomu, aby se staly dalším problémem veřejného zdraví. JAMA J.Am. Med. Doc. 259, 2958–2959. doi: 10.1001 / jama.259.20.2958
CrossRef Full Text / Google Scholar
Saito, h., Yamazaki, h., Matsuoka, h., Matsumoto, K., and Sato, m. (2001). Potenciál související s vizuální událostí u mírné demence Alzheimerova typu. Psychiatrická Klinika. Neurovědci. 55, 365–371. doi: 10.1046 / j. 1440-1819. 2001. 00876.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Schacht, a., Werheid, k., and Sommer, w. (2008). Hodnocení krásy obličeje je rychlé, ale není povinné. Cogne. Ovlivnit. Chovej se. Neurovědci. 8, 132–142. doi: 10.3758 / cabn.8.2.132
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Scott, J. P. R., Mcnaughton, L. R., a Polman, R. C. J. (2006). Účinky deprivace spánku a cvičení na kognitivní, motorický výkon a náladu. Fyziol. Chovej se. 87:408. doi: 10.1016 / j. physbeh.2005.11.009
PubMed Abstract / CrossRef Full Text / Google Scholar
Smallwood, J., Brown, k., Baird, B., and Schooler, J. W. (2011). Spolupráce mezi výchozí režimovou sítí a frontální parietální sítí při výrobě vnitřního myšlenkového proudu. Brain Res.1428, 60-70. doi: 10.1016 / j. brainres.2011.03.072
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Smith, M. E., Mcevoy, L. K., a Gevins, A. (2002). Dopad mírné ztráty spánku na neurofyziologické signály během výkonu úlohy pracovní paměti. Prospal 25, 784-794. doi: 10.1093 / spánek / 25.7.56
CrossRef Plný Text | Google Scholar
Thomas, M. (2003). Neurální základ bdělosti a poruchy kognitivního výkonu během ospalosti ii. účinky 48 a 72 hodin deprivace spánku na probuzení lidské regionální mozkové aktivity. Thalam. Relate. Syst. 2, 199–229. doi: 10.1016/S1472-9288(03)00020-7
CrossRef Plný Text | Google Scholar
Van Dongen, H. (2004). Kumulativní náklady na další bdění: dávkovací účinky na neurobehaviorální funkce a fyziologii spánku z chronické deprivace spánku a celkové deprivace spánku. Prospal 26, 117-126. doi: 10.1016/0739-6260(84)90040-6
CrossRef Plný Text | Google Scholar
Verweij, I. M., Romeijn, N., Smit, D. J., a Giovanni, P. (2014). Deprivace spánku vede ke ztrátě funkční konektivity v čelních oblastech mozku. NEUROSCI BMC. 15:88. doi: 10.1186/1471-2202-15-88
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Wen, X., Liu, Y., Yao, L., and Ding, m. (2013). Top-down regulace aktivity výchozího režimu v prostorové vizuální pozornosti. J. 33, 6444–6453. doi: 10.1523/JNEUROSCI.4939-12.2013
CrossRef Plný Text | Google Scholar
Wiggins, E., Malayka, M., a Kendra, G. (2018). 24-h spánková deprivace narušuje včasnou modulaci pozornosti neurálního zpracování: studie mozkového potenciálu související s událostmi. Neurovědci. Lette. 677, 32–36. doi: 10.1016 / j. neulet.2018.04.022,
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Zhang, L., Shao, Y., Liu, Z., Li, C., Chen, Y., Zhou, Q (2019). Snížená výměna informací pracovní paměti po deprivaci spánku: důkazy z potenciální studie související s událostmi. Před. Neurovědci. 13:408. doi: 10.3389 / fnins.2019.00408
CrossRef Plný Text | Google Scholar
Zhang, Q., Liao, Y., Qi, J, Zhao, Y., Zhu, T., Liu, Z., et al. (2014). Vizuální erp studie inhibice impulzů po zaleplonem indukovaném zdřímnutí po deprivaci spánku. PLoS One 9: e95653. doi: 10.1371 / deník.pone.0095653
PubMed Abstrakt / CrossRef Plný Text / Google Scholar
Leave a Reply